6 Vertebral Column and Turtle Shells
Objectives
- Identify vertebrae from different regions in several clades: sharks, bony fish, salamanders, frogs, turtles, snakes, crocs, birds, mammals.
- Orient vertebrae and identify the major types of vertebral centra.
- Study the morphology of ribs, sternal elements, and hyoid bones across taxa.
- Recognize the elements of the turtle shell in context and the major carapace elements in isolation.
Overview
The vertebrae, in a series, make up the vertebral column, and the vertebral column performs essential functions in support and locomotion. The morphology of the vertebral column evolved in relation to the demands of terrestrial life after a long period of evolution in water, where gravitational demands of body mass on the skeleton are minimized. Regionalization of the vertebral column became more striking as: (1) the demands of terrestrial locomotion favored a more solid attachment between the pelvic girdle and the vertebral column, (2) tetrapods benefited from the ability to move the head independently from the trunk, and (3) the most posterior ribs were lost. Vertebral structures evolved in direct correlation with evolutionary shifts in locomotor pattern, muscle attachment, and environmental exigencies. This lab explores morphological differences of the vertebrae across vertebrates, with an emphasis on the connection between structure and axial position, and the correlation of form and function.
See the terms list here: Terms 6.1–6.2
Axial skeleton: the basics
Overview of vertebral anatomy
Vertebrae articulate with the skull, the ribs, the os coxae, and with each other. Vertebral morphology varies extensively across taxonomic groups, but some features are largely conserved. All vertebrae have a neural arch on the dorsal surface through which the spinal cord extends. The neural arches of a series of vertebrae form a channel called the neural canal. Most caudal vertebrae have a haemal arch on the ventral surface through which blood vessels pass. Both arches can extend into narrow spines, called the neural spine and haemal spine, respectively. The arches and spines provide extra surface area for muscle attachment and lever-arm length for muscle action. Neural and haemal arches rest either directly on the notochord, on an ossified base, or directly on the centrum. The centrum replaces the notochord in many adult vertebrates.
Centra of successive vertebrae articulate with one another to form the vertebral column. A series of apophyses (articular projections) extend from each centrum. Zygapophyses are processes that interlock between successive vertebrae and prevent twisting. Postzygapophyses project caudally from the neural arch and articulate with the prezygapophyses which project cranially from the neural arch. Snakes are especially prone to axial twisting, so they have additional articular surfaces including zygosphenes (projections located cranial to the neural arch and dorsal to the prezygapophyses) that insert into zygantra (caudally oriented pockets on the caudal surface of the neural arch of the preceding vertebra). Fishes have ventral projections called basapophyses for rib articulation. Tetrapods evolved large, lateral projections on their vertebrae that are called transverse processes. Tetrapod ribs articulate with the vertebrae in two locations: the diapophysis, which is a surface on the dorsal end of the transverse process, and the parapophysis, which is on the ventral end of the transverse process. In mammals, the parapophysis is located on the interface between the centra of two vertebrae, and the diapophysis and parapophysis are renamed as the full facet and demifacet respectively.
Overview of rib anatomy
A rib is defined as a rod-shaped structure lateral to the vertebrae. Ribs provide structure to the body wall and attachment sites for trunk and tail muscles. In fishes, each rib has a single head that articulates with the vertebra at the basapophysis. In tetrapods, the ventral ribs were lost and the dorsal limbs evolved into trunk ribs. Trunk ribs have two heads that articulate with the vertebrae. The tuberculum is the dorsal head that articulates with the diapophysis, and the capitulum is the ventral head that articulates with the parapophysis.
Vertebral processes: a reference guide
Diapophysis – the process with which the tuberculum (dorsal head) of the rib articulates. Primitively, the diapophysis was a facet on the neural arch. Diapophyses are found in tetrapods. Parapophysis – the process with which the capitulum (ventral head) of the rib articulates. Primitively, the parapophysis was a facet on the intercentrum. Parapophyses are seen in teleosts (as the basipophysis) and in tetrapods. Prezygapophysis – the process on the cranial portion of the vertebra that articulates with the postzygapophysis of the preceding vertebra. Prezygapophyses are found in some teleosts (modified basal processes) and tetrapods. Postzygapophysis – the process on the caudal portion of the vertebra that articulates with the prezygapophysis of the subsequent vertebra. Postzygapophyses are found in some teleosts (modified basal processes) and tetrapods. Zygosphene – a process that inserts into the zygantrum to provide a secondary articulation with the preceding vertebra; on the cranial surface of the neural arch, complementing the prezygapophysis. Zygosphenes are found in snakes. Zygantrum – a pocket that receives the zygosphene to provide a secondary articulation for the subsequent vertebra on the caudal surface of the neural arch, complementing the postzygapophysis. Zygantra are also found in snakes. Transverse process – any process that sticks out laterally from the vertebra (it is not a very specific term). The diapophyses of amphibians are sometimes referred to as transverse processes, but you should avoid that usage. Transverse processes are exhibited broadly across amniotes.
Types of vertebral centra
Vertebral centra show differences in shape across taxonomic groups, depending on degree and direction of movement and role in support. A quick characterization of centrum morphology is based on the shape of the bony centrum itself, rather than any cartilaginous or fibrous tissue that may adhere to the centrum. Vertebral centra can be assigned to one of five morphotypes:
- Amphicoelous: both ends concave. Common in fishes and early tetrapods.
- Procoelous: anterior end concave, posterior end convex. Common in reptiles.
- Opisthocoelous: anterior end convex, posterior end concave. Seen in Lepisosteus and some salamanders.
- Acoelous (or amphiplatyan): both ends flat. Common in birds and mammals.
- Heterocoelous: both ends shaped like the seat of a saddle, placed transversely at one end and vertically at the other. Common in bird necks.
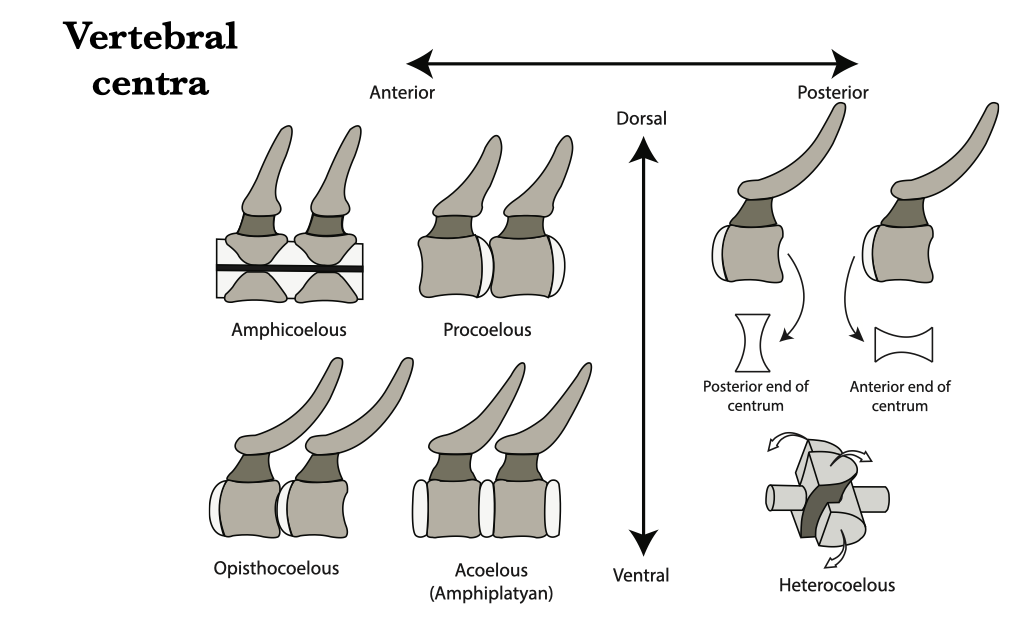
Figure 6.1. Vertebral centra types: amphicoelous, procoelous, opisthocoelous, acoelus, and heterocoelous.
Vertebral evolution
Chondrichthyes & Actinopterygii
The vertebral column of chondrichthyans (sharks, rays, and skates) and actinopterygians (bony fish) are divided into two regions: trunk (the region cranial to the position of the anus) and caudal (the region caudal to the anus). Vertebrae from these regions can be distinguished by the absence (trunk) or presence (caudal) of haemal arches. The head and trunk of organisms in these groups move in conjunction with one another, so these species have no specialized vertebrae near the base of the skull, and therefore no neck.
Chondrichthyan vertebrae are cartilaginous and, in life, the notochord runs through the centrum of each vertebra and through the space between adjacent vertebrae. The vertebrae are amphicoelus. Amia and Lepisosteus have vertebrae that are generally morphologically similar to those of chondrichthyans. Amia are unique, however, in having diplospondyly in the tail, where each caudal vertebra has two centra (see figure of Amia vertebrae). The vertebrae of Lepisosteus have opisthocoelous vertebrae with no canals in the centra for a notochord.
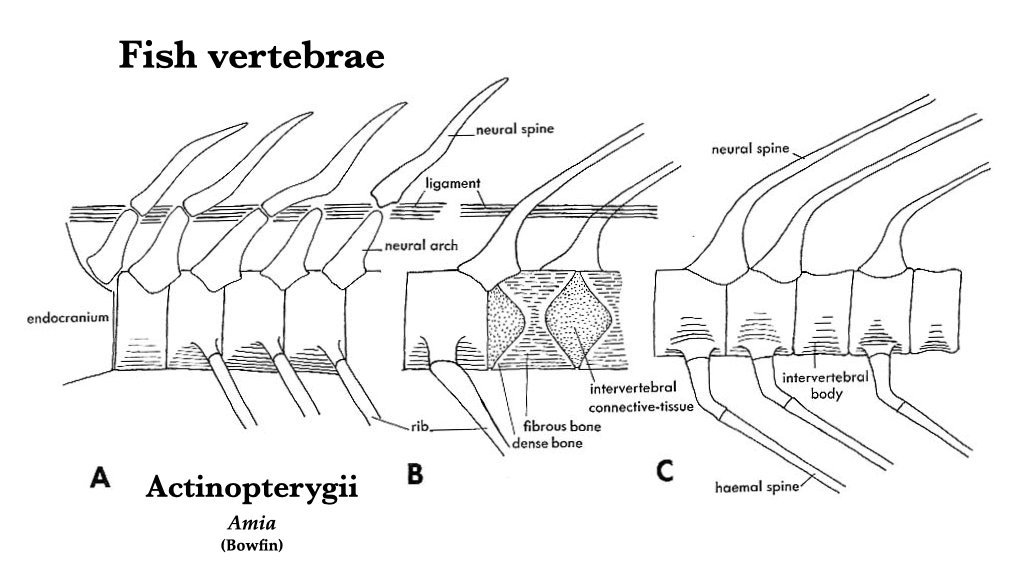
Figure 6.2. Vertebral structure in bowfin (Amia): anterior end of vertebral column, lateral aspect (A); middle trunk vertebrae, lateral (B); and caudal vertebrae showing transition to diplospondyly, lateral (C).
The vertebrae of teleost actinopterygians have amphicoelous centra. Each neural and haemal arch terminates in a long, sharp spine, all of which are directed caudally. The trunk vertebrae have paired projections on the lateral sides of the centrum which articulate with long, thin ribs. The transition between trunk and caudal vertebrae can best be observed on a complete skeleton where, moving from cranial to caudal, the basapophyses become longer and the ribs become shorter. The tail begins where the ribs are no longer present and the basapophyses fuse to form haemal arches.
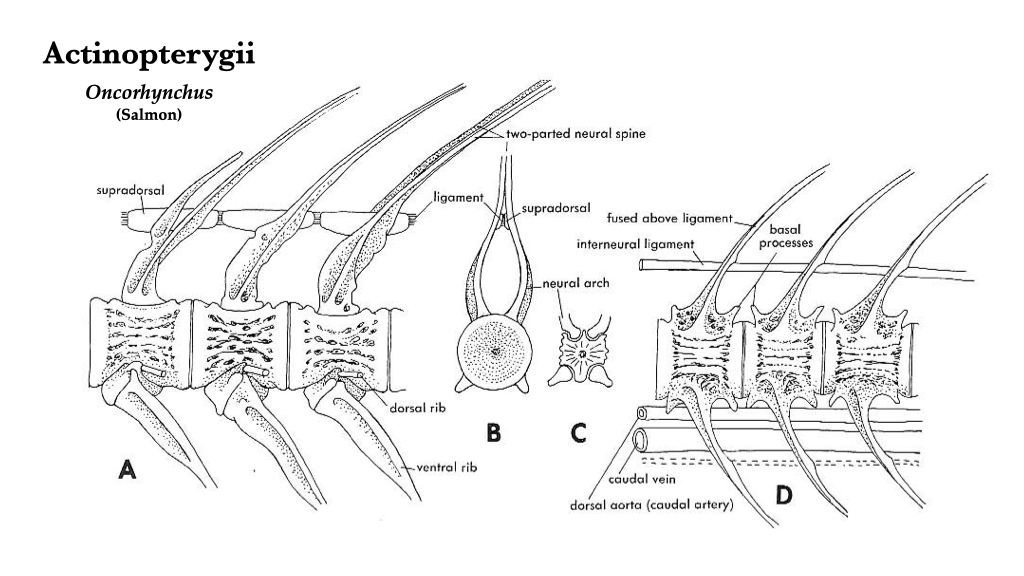
Figure 6.3. Vertebral structure in teleosts, as exemplified by salmon (Oncorhynchus): A, trunk vertebrae, lateral view; B, trunk vertebrae, cranial view; C, trunk vertebrae, caudal view; D, caudal vertebrae, lateral view.
Amphibia
Amphibians, most of which spend at least some time out of the water, have a more differentiated spinal column than do fish. In addition to trunk and caudal vertebrae, amphibians generally have a single cervical vertebra that articulates with the skull, and a single sacral vertebra that contacts the os coxae. The pre- and postzygapophyses help provide strong, stable intervertebral articulations for support of the body outside the water, and transverse processes provide attachment for ribs and muscles.
Caudata (Salamanders)
Five regions of the vertebral column can be recognized in salamanders: cervical (a single vertebra), trunk (usually bearing ribs), sacral (attached to the os coxae), caudosacral (two to four transitional vertebrae, with the first being immediately posterior to the sacral vertebra, and the last being the first in the vertebral column to exhibit a well-developed haemal arch), and caudal (with haemal arches that are caudal to the caudosacral vertebrae). Centra are usually amphicoelous (except in salamandrids and some plethodontids, which are opisthocoelous). Note that the tuberculum interglenoideum is a process on the atlas that projects into the foramen magnum, and it is unique to salamanders.
Anura (Frogs)
In order to improve structural stability and force transfer, frogs have reduced the number of vertebrae and only have three vertebral regions. As with other amphibians, a single cervical vertebra is present, but there are four to eight (usually seven) trunk vertebrae, and a single sacral. Frogs lack a tail, instead they have an elongate rod composed of fused post-sacral vertebrae, the urostyle. The urostyle articulates cranially with the sacral and caudally with the pelvis. In basal frog lineages, the adults have free ribs, but otherwise frog vertebrae have well-developed transverse processes. The neural arches are low, with small zygapophyses. Most species, including Rana, have procoelous centra, but opisthocoelous centra occur in some.
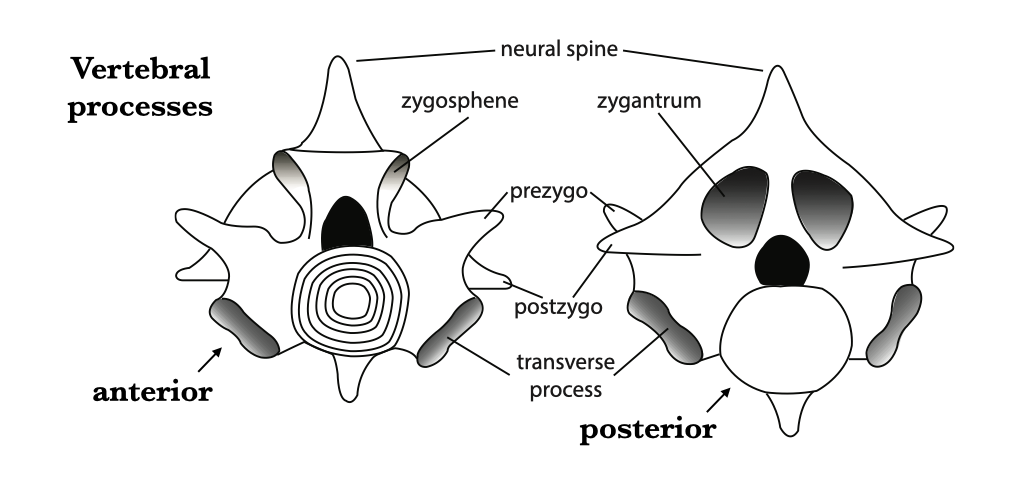
Figure 6.4. Vertebral processes of an idealized vertebra in anterior and posterior views.
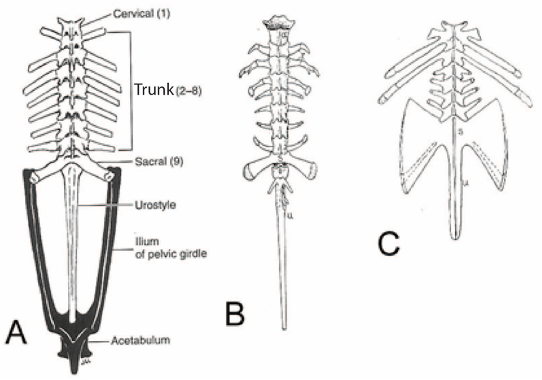
Figure 6.5. Morphological diversity of the vertebral column in Anura. Superior views of generic anuran (A), Ascaphus (B), and Pipa (C). Illustration of the generic anuran from Kent and Miller (1997) under fair use guidelines; Ascaphus and Pipa from Kingsley (1925) under CC0 public domain.
Gymnophiona (Caecilians)
Caecilians have a single cervical vertebra and 60 to 285 trunk vertebrae, no sacral vertebrae (caecilians are legless), and few, irregularly shaped caudal vertebrae. Most of the trunk vertebrae have bicipital (two-headed) ribs and high neural arches, but they lack haemal arches although there may be anomalous, haemal arch-like structures in the cloacal region. The rib-bearing vertebrae are especially distinctive because the diapophyses are positioned on the anterior part of the neural arch, and the parapophyses are on the extreme anterior part of the centrum.
Amniotes
The cervical region of amniotes is more extensively modified for the support and movement of the skull. The atlas and axis become specialized in amniotes for this purpose. The axis usually has an anterior projection — the odontoid process or dens — that extends through the anteroposteriorly compressed atlas and into the foramen magnum. The dens permits movement of the head from side to side, which is constrained in fishes and amphibians. In some taxa (e.g., crocodilians) a proatlas is present anterior and dorsal to the atlas.

Figure 6.6. Skeleton of a snake in superior view. Cottonmouth (Agkistrodon piscivorus) specimen from the UCB teaching collection. Cottonmouth photo from Flickr user Peter Paplanus under CC BY 2.0.
Testudines
In turtles, there are eighteen presacral vertebrae, and the anterior eight of these are cervicals. There are two major groups of turtles, Cryptodira and Pleurodira, and they are distinguished in part by the way they fold their head into the shell. Generally, cryptodires fold the neck in a vertical plane, pleurodires in a horizontal plane. The atlas of cryptodires is usually in three parts, while that of pleurodires is usually fused. The trunk vertebrae are involved in the formation of the carapace (upper part of the shell), with each vertebra corresponding to a bony plate at midline. Neural arches are displaced forward to an intercentral position. The ribs are laterally expanded to form part of the costal portion of the carapace. The sacrals are usually the 19th and 20th vertebrae. The caudals are usually small, few, and procoelous.
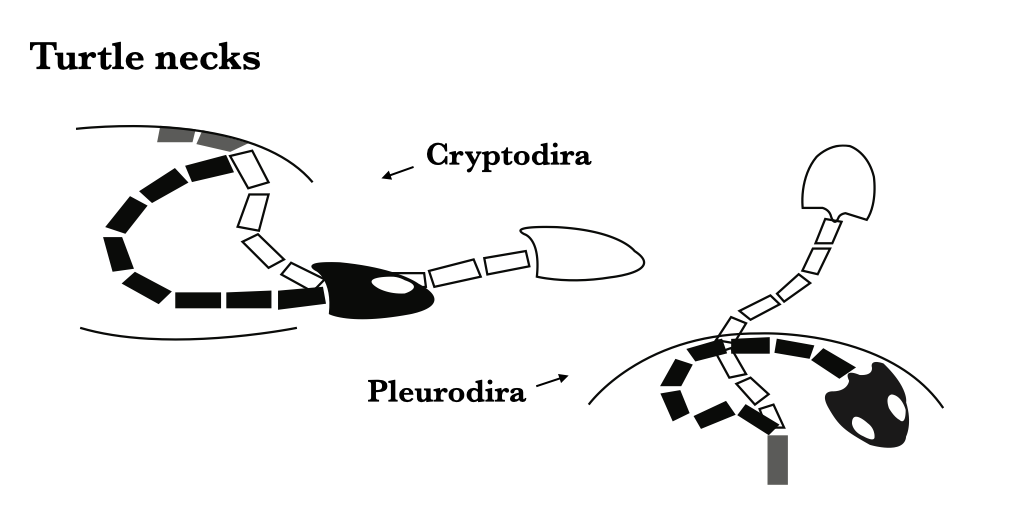
Figure 6.7. Turtle necks: cryptodirans bend the neck dorsoventrally/vertically to retract the head (left), pleurodirans bend the neck sideways/horizontally to retract the head (right).
Squamata
The Squamata includes amphisbaenians, “lizards,” and snakes. The majority of lizards have procoelous centra, although there is a lot of variation. Lizards have cervical, trunk, sacral, and caudal vertebrae. Many lizards have eight cervical vertebrae. The first cervical vertebra articulates with the cranium and the first trunk vertebra has ribs that connect to the sternum. Squamates have two sacral vertebrae that articulate with the pelvis. Caudal vertebrae of almost all squamates (if not evolutionarily lost) have autotomic septa, a transverse plane that bisects the vertebra and facilitates breakage, allowing the tail to break off when the lizard is threatened and needs to escape. The tail can regrow if it breaks at the septum, but the new tail has no vertebrae and is composed of a cartilaginous rod. The vertebral column of snakes is not easily divided into recognizable regions. The vertebral column plays a key role in snake locomotion, and as a result, the vertebrae of most snakes are complex. The atlas/axis complex is much like that in other lizards, but there is no reliable way to distinguish the other cervicals from the trunk vertebrae. Zygosphenes and zygantra are present.
Crocodylia
Crocodilians have cervical, thoracic, lumbar, sacral, and caudal regions in the vertebral column. There are 9 cervical vertebrae, each of which has cervical ribs that do not attach to the sternum. The crocodylian atlas is unique in that it has 2 neural arches, a ventrally placed intercentrum, and a dorsally placed proatlas. The centrum of the atlas is fused with the axis and contributes to the dens. Cervical rib length increases posteriorly. The trunk vertebrae are differentiated into thoracic vertebrae that have ribs, and lumbar vertebrae that do not have ribs. There are 2 sacral vertebrae in living crocodilians, both of which have ribs that articulate with the os coxae. Although most of the vertebral centra are procoelous, the centrum of the first postsacral vertebra (the caudosacral) is biconvex, lacks a haemal arch, and is considered transitional between the sacral and caudal regions. The anterior caudal vertebrae have haemal arches and well-developed transverse processes which taper off posteriorly.
Aves
The vertebral column of birds is characterized by increased rigidity in all but the cervical region. The atlas is ring-shaped and lacks a centrum. The cervical vertebrae either lack ribs or the ribs do not connect to the sternum. The extreme mobility of the avian neck is partly a function of the heterocoelous centra. The thoracic vertebrae bear ribs that contact the sternum, and the vertebrae are often fused to each other. The thoracic ribs have a posteriorly directed uncinate process that strengthens the rib cage and provides muscle attachment sites. The lumbar, sacral, and some caudal vertebrae are fused to each other and fused to the os coxa to form the synsacrum. There are usually a few free caudal vertebrae posterior to the synsacrum. The vertebral column terminates in the pygostyle, a structure formed by the fusion of several caudal vertebrae.
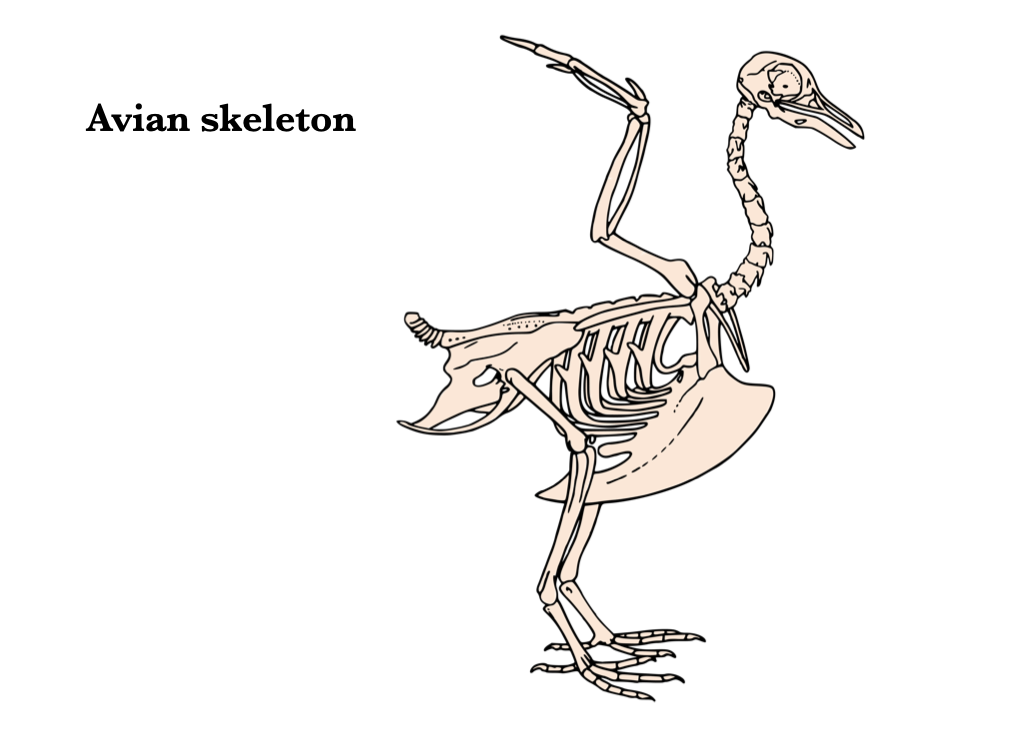
Figure 6.8. Avian skeleton from Wikipedia user mario modesto under CC BY 2.5, based on the original illustration from BIODIDAC.
Mammalia
Mammals have cervical, thoracic, lumbar, sacral, and caudal vertebrae. All living mammals, with the exception of sloths and manatees, have seven cervical vertebrae. The atlas is ring-shaped, as in birds, but it has wing-like transverse processes that are derived from cervical ribs. Most other cervical vertebrae have transverse foramina, canals for vertebral arteries and veins. Most mammalian cervical vertebrae are acoelous (amphiplatyan). On the thoracic vertebrae, diapophyses (full facet) on the transverse processes articulate with the tubercles of the ribs, and parapophyses (demifacet) on the centra articulate with the capitula of the ribs. True ribs articulate with the sternum, while false (“floating”) ribs are those that join the preceding rib or are unattached ventrally. The lumbar vertebrae have long, anteriorly directed transverse processes. The anterior lumbar vertebrae may have mammillary processes dorsolateral to the prezygapophyses. The sacral vertebrae of mammals are fused into a sacrum, which articulates with the ilia.
Turtle Shells
The turtle shell is composed of two primary regions: the carapace (upper shell) and the plastron (lower shell). Both carapace and plastron are overlain by epidermal scutes. These two shell regions are fused along the lateral sides of the body by “bridges.”
Carapace
The turtle carapace is made up of five bony element types. Neurals are the plates that run down the midline and are formed from modified neural arches. The neural canal runs immediately ventral to the neural plates, with the vertebrae attached below. The neurals are symmetrical, which facilitates their identification. Costals are modified ribs, fused and expanded to form most of the area of the carapace, lateral to the neurals. Costals are identifiable by their elongate form, and by the protruding head of the rib on the ventral side of the proximal end, which articulates with the vertebrae, as in other vertebrates. Peripherals are the plates around the border of the shell, which generally have a smooth, beveled edge which lacks the sinuous suture pattern that is evident on all other plates of the carapace.
Finally, there are two specialized plates at the anterior and posterior ends of the shell. The nuchal is the anteriormost plate, where the line of neurals meets the circle of peripherals. It is recognizable for the distinctive pattern of lines from the overlying epidermal scutes, and for its bilateral symmetry. The pygal is the posteriormost plate of the carapace, where the posterior end of the line of neurals meets the circle of peripherals. The pygal has bilateral symmetry and a distinctive pattern of scute lines. There is also a small suprapygal between the last neural and the pygal.
Plastron
The plastron has fewer elements than the carapace. The epiplastron is the anteriormost bony element; it is paired, and generally curved around the entoplastron (the only unpaired element of the plastron). The hyoplastron includes the anterior portion of the bridge which connects the plastron and the carapace. The hypoplastron includes the posterior portion of the bridge. Finally, the xiphiplastron composes the posteriormost part of the plastron.
Hard-shelled and soft-shelled turtles
Soft-shelled turtles encompass the families Trionychidae and Carettochelyidae, while hard-shelled turtles comprise the remainder. Soft-shelled turtles generally have reduced the carapace and plastron, often leaving spaces in the dome of the carapace between the costals, although this sort of reduction also occurs in some hard-shelled turtles, such as sea turtles.
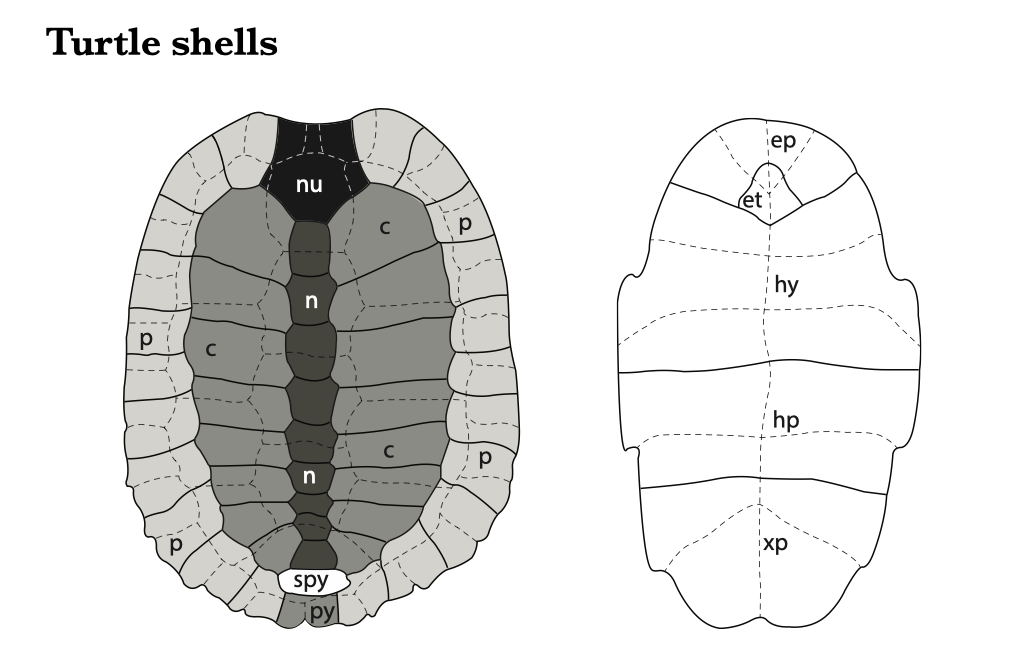
Figure 6.9. Turtle shell bones of the carapace (left) and plastron (right). Superimposed dashed lines represent the removed scutes. Abbreviations: nu (nuchal), n (neural), p (peripheral), c (costal), py (pygal), spy (suprapygal), ep (epiplastron), et (entoplastron), hy (hyoplastron), hp (hypoplastron), xp (xiphiplastron).



{kind=link}