3 Amphibian and Reptile Skulls
Objectives
- Identify all of the major elements of the non-mammalian tetrapod skull.
- Recognize the development origin (endochondral or intramembranous) and cranial region (neurocranium, splanchnocranium, dermatocranium) to which each object belongs.
- Observe the differences in morphology among species from different taxonomic groups and under divergent functional demands.
Overview
In this lab, we transition from fish skulls to tetrapod skulls. The evolutionary transition of vertebrates from water to land was accompanied by large changes in the demands of feeding, gas exchange, and sensation. Adapting to such large changes in the environment involved major shifts in morphology. By walking through the evolutionary history of vertebrate skulls, you will learn to recognize morphological differences among taxa based on both their ancestry and their functional demands. After this lab, you will be able to identify the objects listed below in bold face in the articulated skull, and you will learn to identify the objects with asterisks (*) in isolation. Bones are lost, fused, and gained throughout evolutionary history, and we have provided separate lists for each taxonomic group. The diagrams are organized phylogenetically, and the list of specified objects for a group can be found at the beginning of the section containing the diagram for that group. For a given clade, you only need to know the objects in the pertinent list.
See the terms list here: Terms 3.1–3.6
Strategies for success
The table on the following page sorts the bones by cranial region as well as by their location or by the structure to which they contribute. The dermal bones tend to be faithful to their position relative to the nares, orbits, and fenestrae, and you might start with a focus on those. Try to develop a consistent approach to any skull that you examine. You might look for the nasal bones, follow those to the frontals, and continue from there. Or perhaps you will want to start with the lower jaw and dispense with that before continuing to the cranium. Try to be systematic and creative in your approach to learning skull morphology.
Checklist of skull bones by region and evolutionary/developmental origin
|
Skull region |
Neurocranium |
Splanchnocranium |
Dermatocranium |
|
Braincase: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Skull Roof: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Circumorbital: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Temporal: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Marginal Jaw: |
|
|
|
|
|
|
|
|
|
Palatal: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Lower Jaw: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Early Amphibian Skulls
The evolutionary transition from water to land was marked by major changes in the skull and skeleton of the early tetrapods. The more derived skulls tend to be longer, due mostly to the elongation of the snout that correlate with differences in feeding. Many skull bones underwent fusion, and some joints became akinetic. The evolution of internal nostrils, or choanae, was a means to pass air from outside the body into the newly evolved lungs for respiration. With the dependence on lungs, gills were lost and the opercular series was lost with them. Eusthenopteron, Ichthyostega, and Seymouria are all early branches from the lineage that led to modern tetrapods. Eusthenopteron had foot-like fins, Ichthyostega had fin-like feet, and Seymouria probably had the familiar five-toed feet of many modern tetrapods.
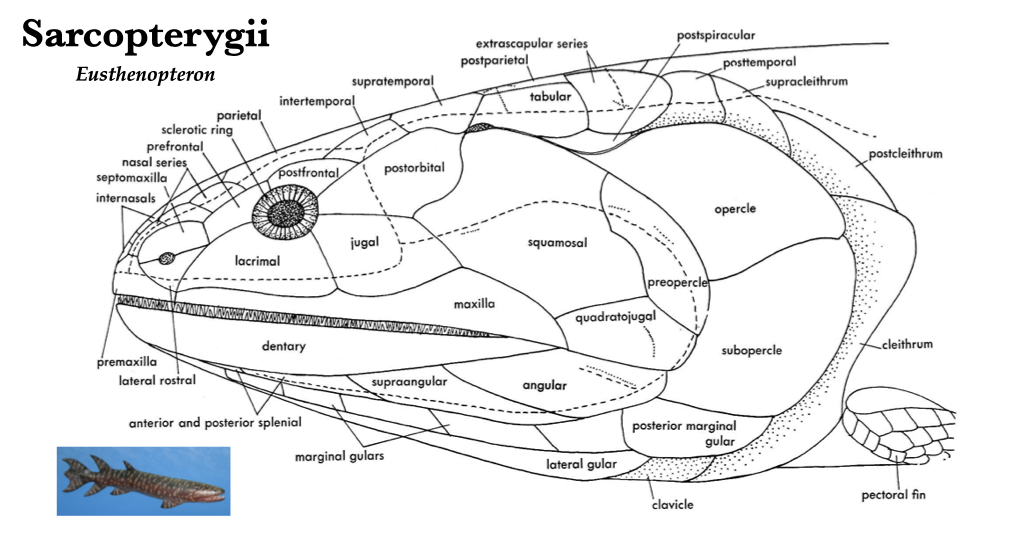
Figure 3.1. Skull of Eusthenopteron, a prehistoric sarcopterygian fish closely related to tetrapods, lateral view. From Jollie (1962) under CC0 public domain; Eusthenopteron reconstruction from Wikipedia user Dr. Günter Bechly under CC BY-SA 3.0.
{kind=link}
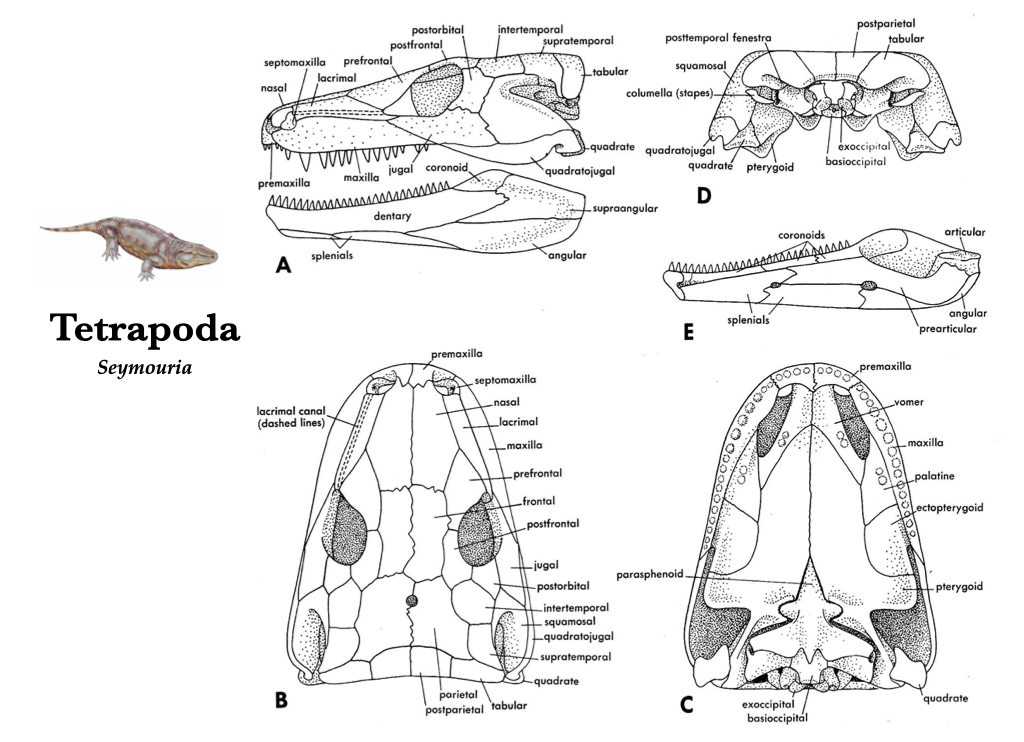
Figure 3.2. Skull of Seymouria, a reptile-like tetrapod from the Early Permian in lateral (A), dorsal (B), ventral (C), posterior (D) views, and medial view of the right lower jaw (E). Illustrations from Jollie (1962) under CC0 public domain; Seymouria reconstruction from Wikipedia user Dmitry Bogdanov under CC BY 3.0
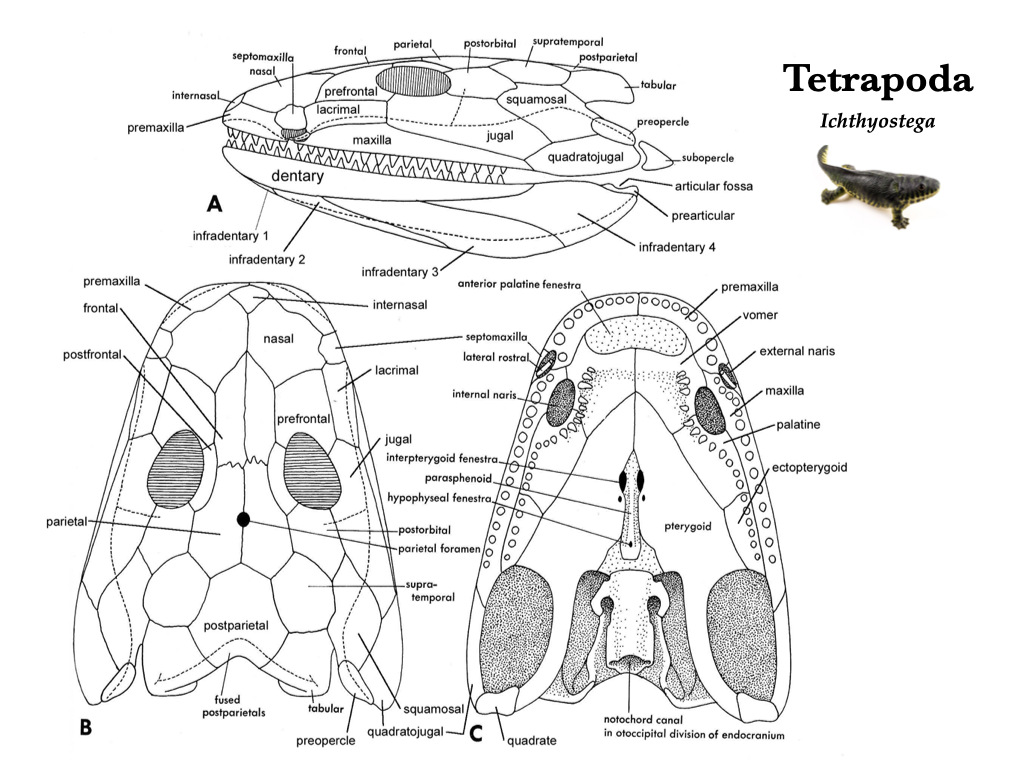
Figure 3.3. Skull of Ichthyostega, a basal tetrapod, in lateral (A), dorsal (B), and ventral (C) views. Illustrations from Jollie (1962) under CC0 public domain: Ichthyostega reconstruction on right from Wikipedia user Matteo De Stefano/MUSE under CC BY-SA 3.0.
{kind=link}
Lissamphibian Skulls
The skulls of modern amphibians (Lissamphibia) are generally simplified relative to those of their ancestors. This includes a great reduction in the number of bones present in the skull through losses and fusions. The three major groups of lissamphibians (Gymnophiona, Caudata, Anura) share similarities in bone reduction and fenestration pattern, including the loss of the supratemporals, intertemporals, tabular, postparietals, jugals, and postorbitals. The teeth of lissamphibians are present on the premaxilla, maxilla, vomer, and sometimes on the palatine. The skulls have relatively short occipital regions, and all members have paired occipital condyles. The basioccipital and supraoccipital bones are not ossified. The prootic and opisthotic may be fused to the exoccipitals. The ventral chondrocranium is covered by the dermal parasphenoid, and the quadrate is fused to the otic region. Although there exist many deep similarities among them, the three major groups of lissamphibian look quite dissimilar overall!
Caecilian skulls (Gymnophiona)
Caecilians are a lineage of largely fossorial lissamphibians. Their skulls reflect the digging habit through the loss and fusion of many bone elements. Caecilian skulls are heavily ossified and exceptionally compact. Caecilians have a sphenethmoid bone and an os basale, both of which are formed through fusion of many other bones. They have reduced eyes, sometimes with bone obscuring the orbits completely. Due to the heavy ossification, caecilians have evolved a unique jaw closure mechanism in which the interhyoideus muscle replaces the adductor mandibulae as the primary muscle of jaw closure, thus converting the system from a third- to a first-class lever. Caecilians have a very large retroarticular process that provides a long in-lever.
Salamander skulls (Caudata)
Generally, salamander skulls are relatively broad and flat. Salamander skulls are generally used for feeding and breathing on land, and sheets of muscle attach to the broad surfaces of the skull bones. The frontal and parietal bones remain unfused across the midline in salamanders. Salamanders have teeth on the vomers. How these teeth are arranged is unique by family, so one is actually able to identify the family to which a salamander skull belongs based on its vomerine dental pattern. All salamanders and all frogs lack ectopterygoids, postfrontals, and the basisphenoid. Supraoccipital and basioccipital bones are lost in all salamanders, and palatines are lost during metamorphosis in most taxa. Depending on the family of a salamander, the skull may be missing premaxillae, nasals, maxillae, septomaxillae, lacrimals, or prefrontals.
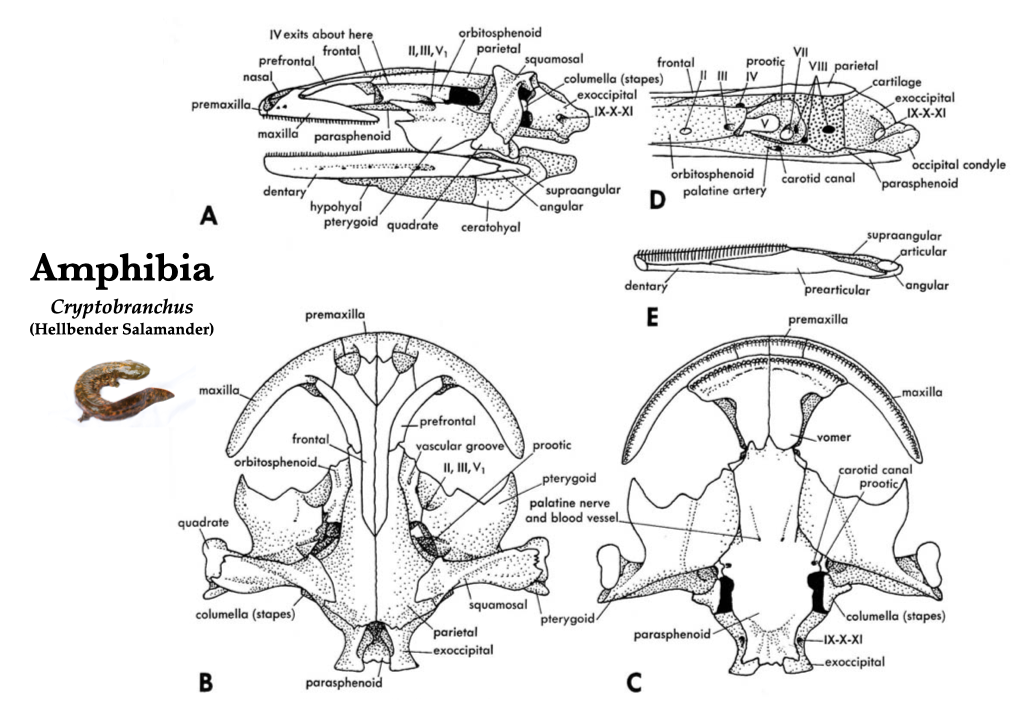
Figure 3.4. Skull of a hellbender salamander (Cryptobranchus) in lateral (A), dorsal (B), ventral (C), and medial (D) views, and right mandible in medial view (E). Illustrations from Jollie (1962) under CC0 public domain; Cryptobranchus reconstruction on left from Flickr user Brian Gratwicke under CC BY 2.0.
Frog skulls (Anura)
With the exceptions of the sphenethmoid, exoccipitals, and prootics, most of the neurorocranium does not ossify in anurans. The choanae are surrounded by the nasal bones dorsally and by the toothed vomers ventrally. The premaxillae and maxillae also have teeth. Anurans, unlike other amphibians, have a quadratojugal that connects the maxilla with the cranium. The mandible of anurans is unique in possessing an anterior cartilaginous bone called the mentomeckelian, and the articular is not ossified. The frontals and parietals fuse to form a frontoparietal. All salamanders and all frogs lack ectopterygoids, postfrontals, and the basisphenoid. All frogs lack the prefrontal, postfrontal, lacrimal, postorbital, jugal, tabular, supratemporal, supraoccipital, basioccipital, and ectopterygoid bones.
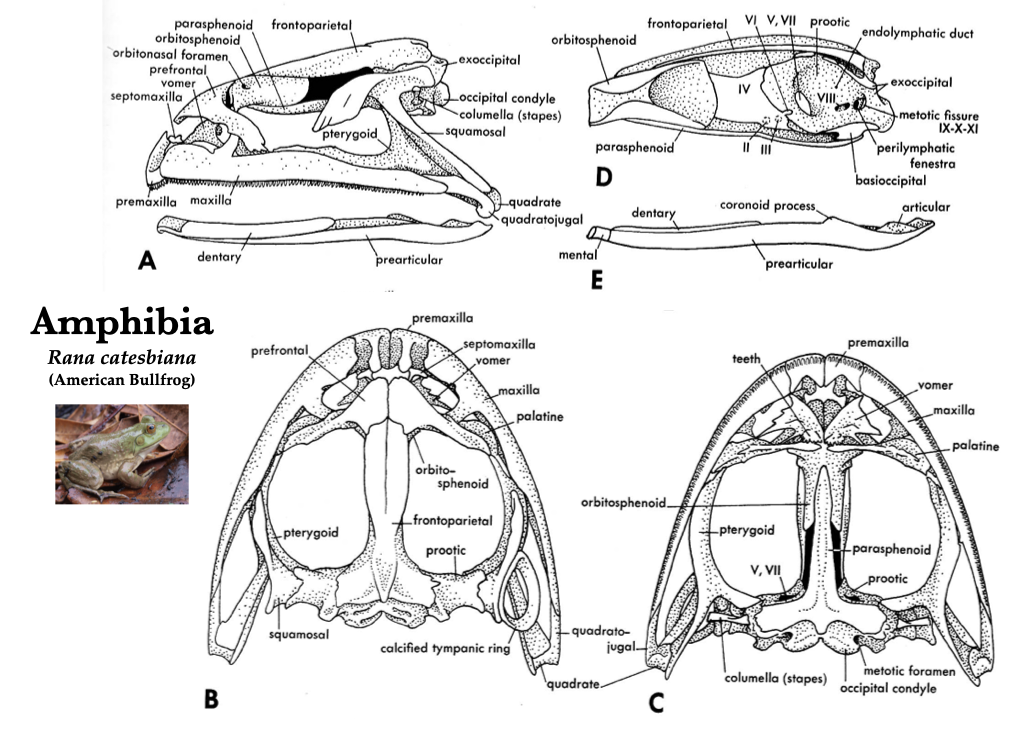
Figure 3.5. Skull of a bullfrog (Rana catesbeiana) in lateral (A), dorsal (B), ventral (C), medial (D) views, and medial view of right mandible (E). Illustrations from Jollie (1962) under CC0 public domain; Rana catesbeiana photo from Flickr user Will Brown under CC BY 2.0.
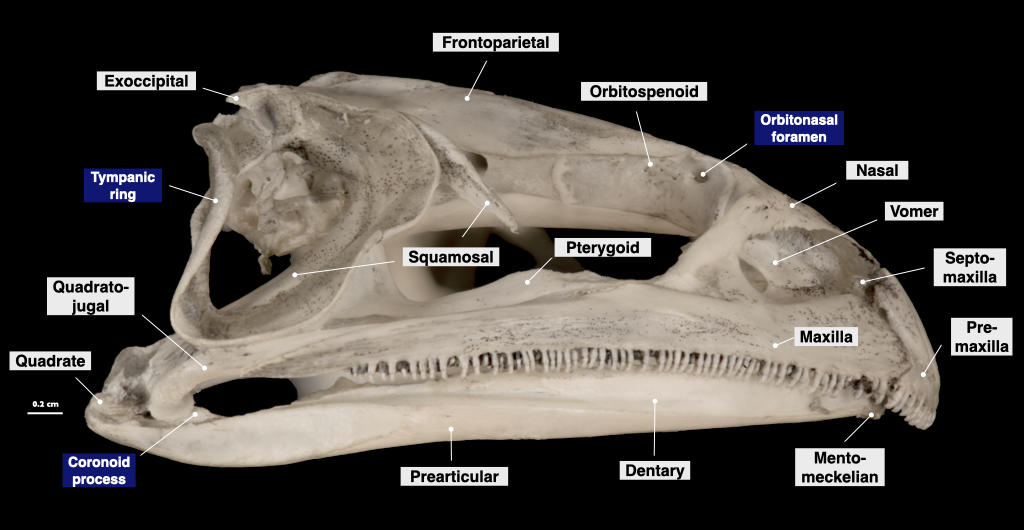
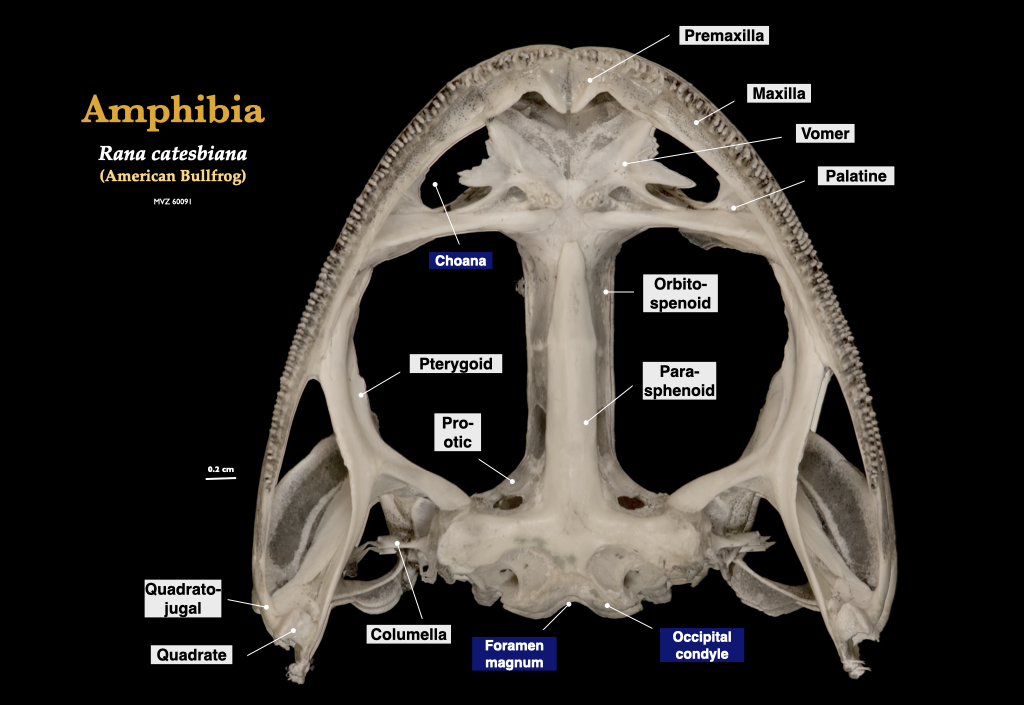
Figure 3.6. Skull of a bullfrog (Rana catesbeiana, MVZ 60091) in lateral view (top) and cranium in inferior view (bottom).
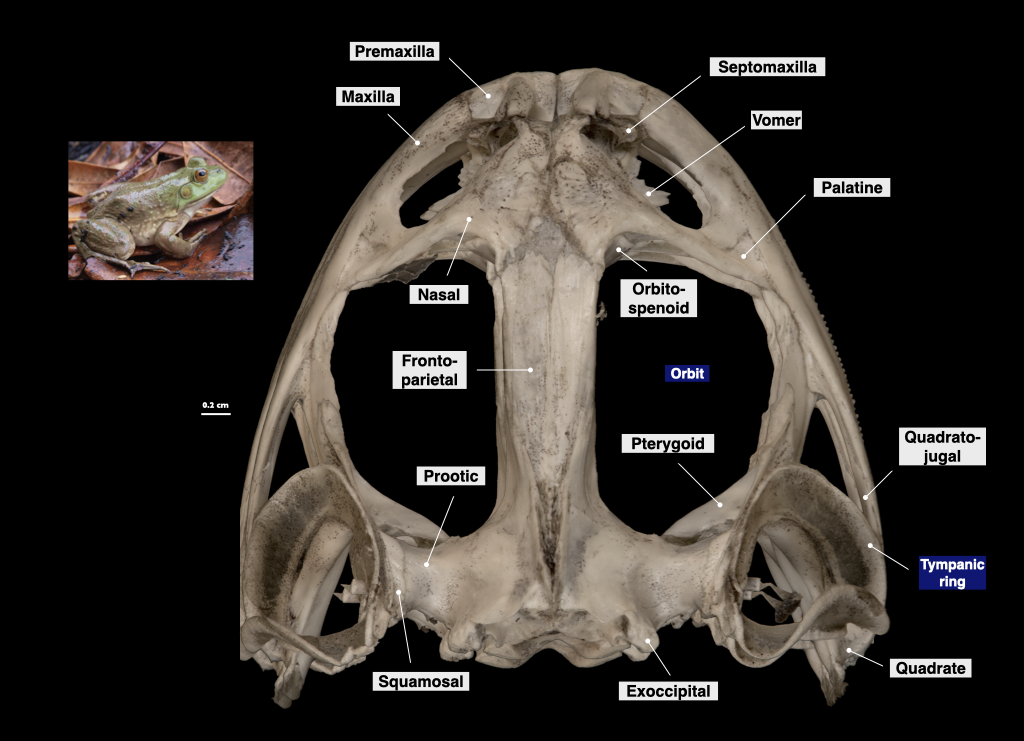
Figure 3.7. Cranium of a bullfrog in superior view (Rana catesbeiana, MVZ 60091).
Basal Amniote Skulls
The primitive amniote skull is typically represented by a generalized “reptile” skull. Amniote skulls are usually well-ossified and are much less reduced than lissamphibian skulls. Basal amniotes have a single occipital condyle formed by contributions from the basioccipital and (usually) the exoccipitals. The prootics are ossified and usually separate from the rest of the otic region. The frontals and parietals are frequently fused to form a single frontal and a single parietal. The interparietal foramen is frequently present, and the sphenethmoid is lost completely.
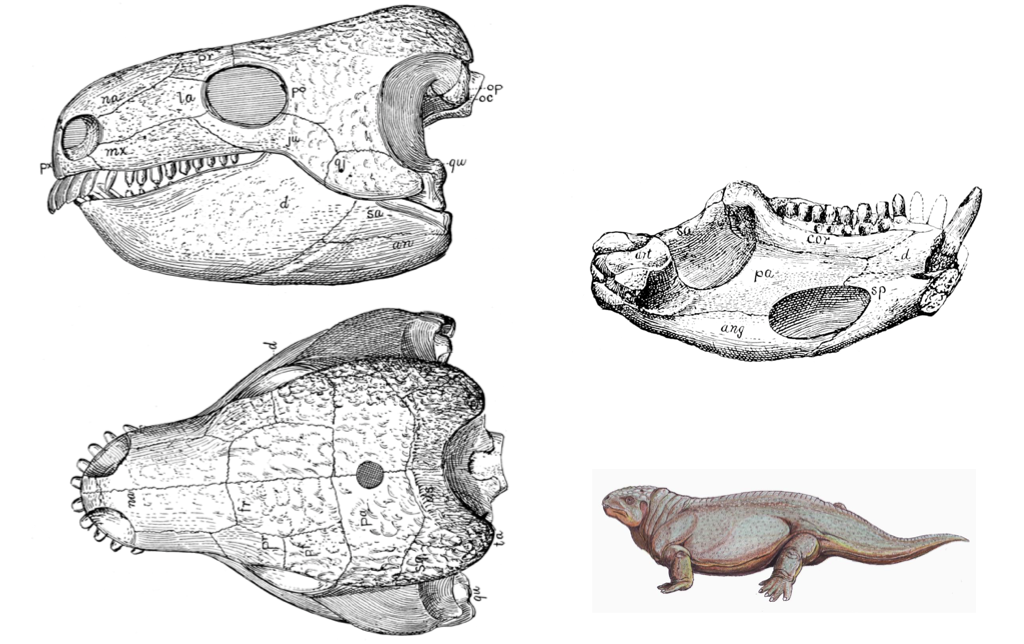
Figure 3.8. Basal amniote skull (Diadectes) from Williston and Gregory (1925).
at = articularan = angularcor = coronoidd = dentaryds = postparietalfr = frontalju = jugalmx = maxillana = nasaloc = occipital condylepa (dorsal view) = parietalpa (on mandible) = prearticularpf = postfrontalpo = postorbitalpr = prefrontalpx = premaxillaqj = quadratojugalqu = quadratesa = surangularsp = splenialsq = squamosalta = tabular
Squamate Skulls (Squamata)
The skulls of amphisbaenians, “lizards,” and snakes (Squamata) have a relatively high amount of mobility or cranial kinesis, and all taxa except chameleons have a moveable joint between the quadrate and the squamosal. The quadratojugal is absent. All four objects of the occipital complex (basioccipital, left exoccipital, right exoccipital, supraoccipital) participate in the foramen magnum. We discuss lizards and snakes separately because their skulls are so divergent.
“Lizard” skulls
Lizard skulls have as many as four moveable joints, permitting a high amount of cranial kinesis. These joints change the geometry of the skull as they move allowing lizards to redistribute bite forces to improve prey capture. The premaxillae and maxillae typically house the teeth, but teeth may also be located on the pterygoids and palatines in some groups, but there are no teeth on the vomer. Each mandible is composed of a dentary, coronoid, angular, surangular, articular, and splenial, and a bony symphysis connects the two mandibles.
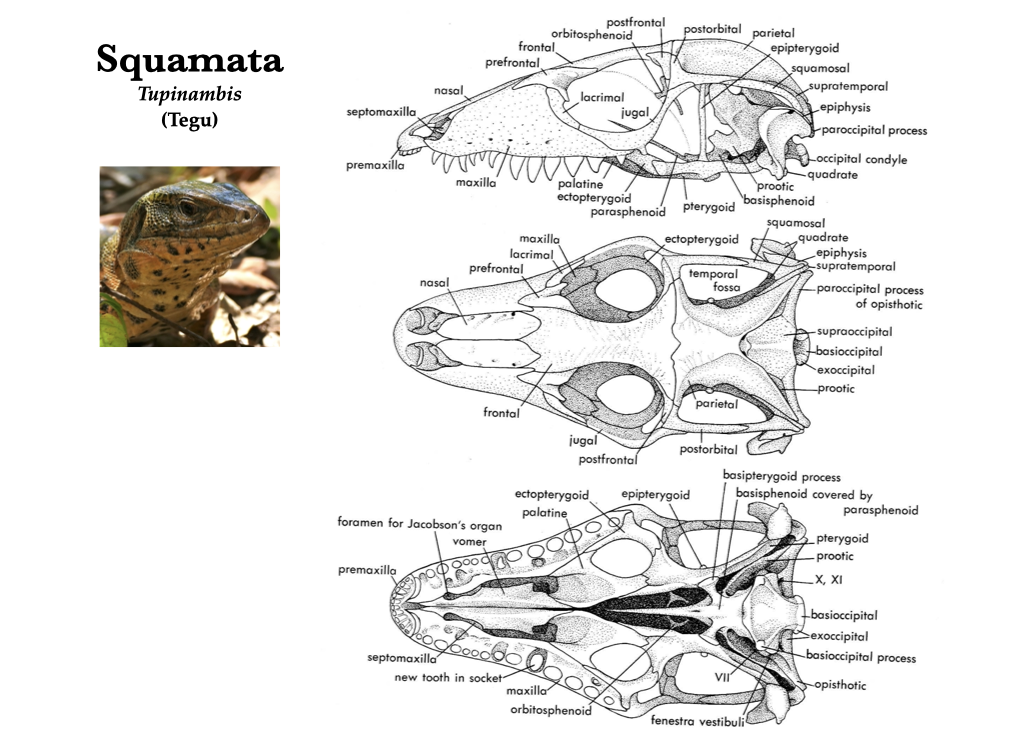
Figure 3.9. Skull of a teiid lizard (Tupinambis) in lateral, dorsal, and ventral views (top to bottom). Illustrations from Jollie (1962) under CC0 public domain; Tupinambis photo from Wikipedia user Bernard DUPONT under CC BY-SA 2.0.
_(28688565543).jpg){kind=link}
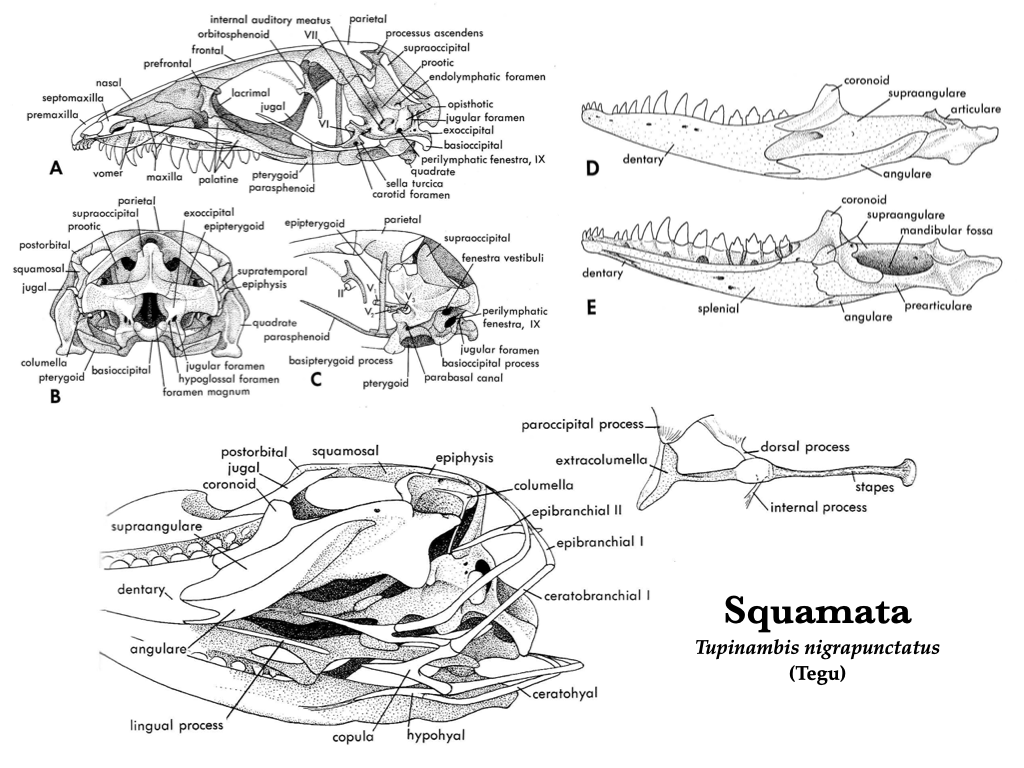
Figure 3.10. More views of the skull of Tupinambis (Teiidae). Illustrations from Jollie (1962) under CC0 public domain.
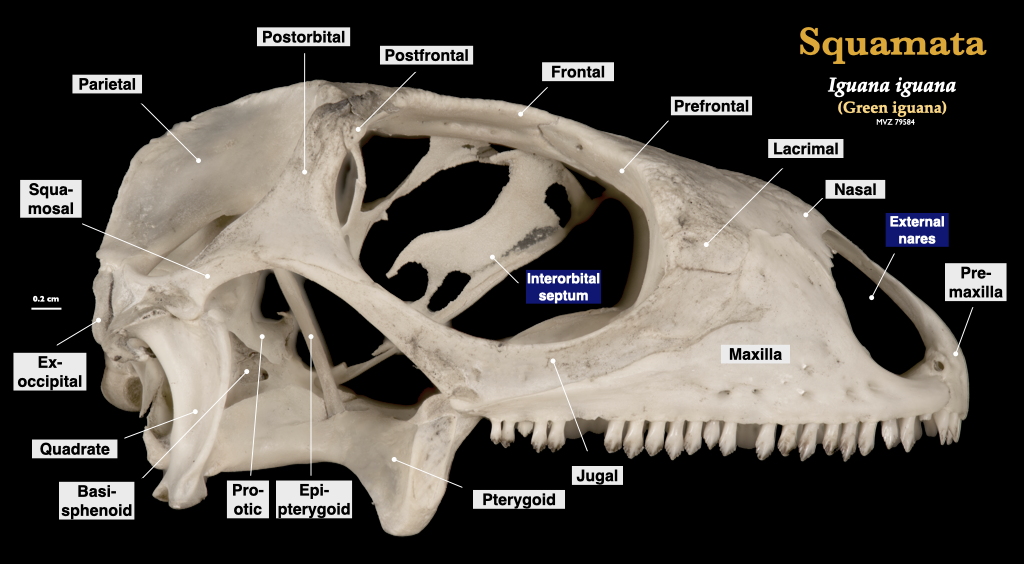
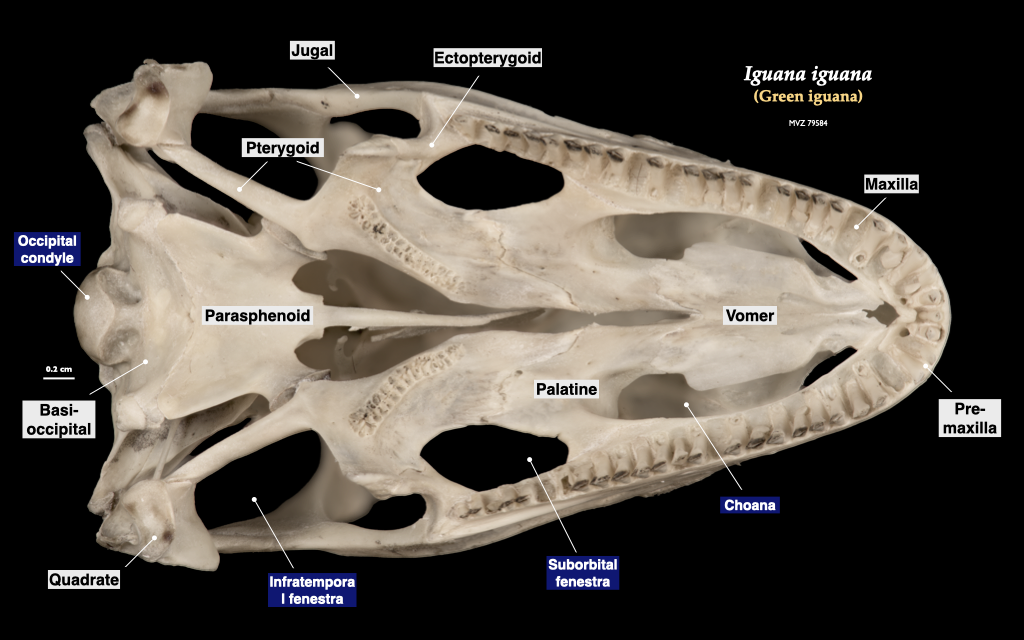
Figure 3.11. Cranium of an iguana (Iguana iguana, MVZ 79584) in lateral (top) and inferior (bottom) views.
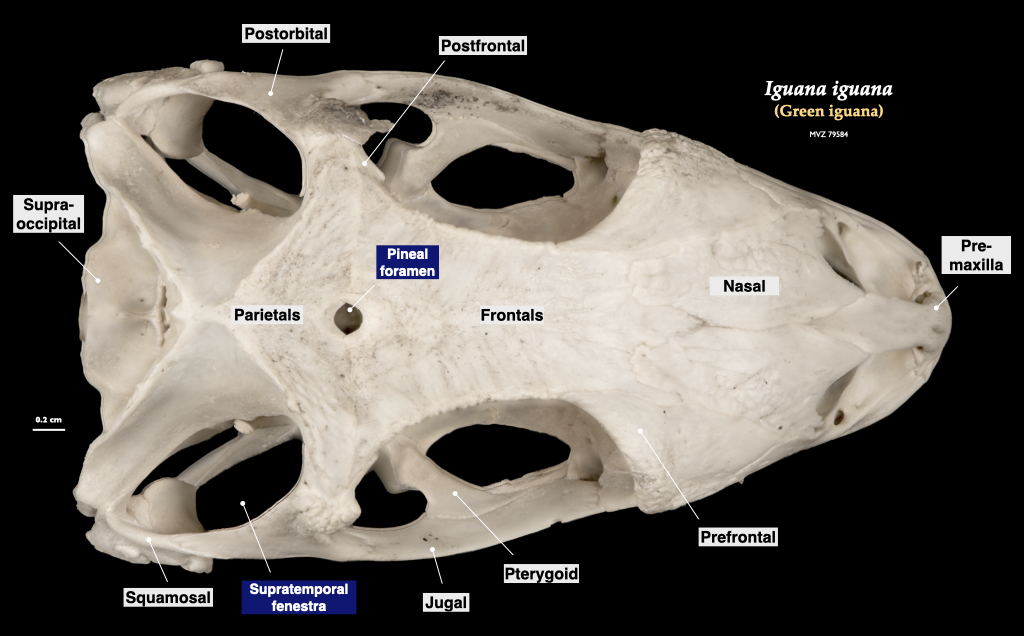
Figure 3.12. Skull of an iguana in superior view (Iguana iguana, MVZ 79584).
Snake skulls (Serpentes)
Snakes are a highly derived group of squamates that likely descend from a fossorial ancestor. Most snakes have evolved to consume prey that are relatively large, and this involved extensive modification of the skull to accommodate a novel feeding mechanism. Snake skulls have less fusion and more loose connections between bones. Snake mandibles are attached to each other by a ligamentous attachment only, and this allows the mouth to stretch around large prey. The bones that make up the palate are usually not fused to the skull but are instead attached by ligaments. This includes the toothed pterygoids. These mobile bones are used to “walk” the snake’s mouth around the prey item. Mobile joints are also present between the frontals, nasals, and prefrontals. Venomous snakes have a particularly small premaxilla that lacks teeth. The maxilla is also small, but it has fangs that can be rotated anteriorly for venom injection. Snakes lack lacrimal, squamosal, epipterygoid, jugal, and quadratojugal bones.
Turtle Skulls (Testudines)
Turtle skulls have a complete covering of dermal bones and, instead of teeth, they have keratinous sheaths covering the upper and lower jaws. All three elements of the turtle otic capsule are ossified, but the epiotic and opisthotic are united with the supraoccipitals, whereas the prootic is not. In some primitive turtles, a false roof composed of the postfrontals, parietals and squamosals is elevated atop the true cranial roof. Turtles have sclerotic ossicles around the eyes. Turtles do not have nasals, parasphenoid, postfrontal, postparietal, or ectopterygoid bones. They usually lack the lacrimals, and the premaxillae and palatines are relatively small. As many as six bones fuse to form the turtle mandible.
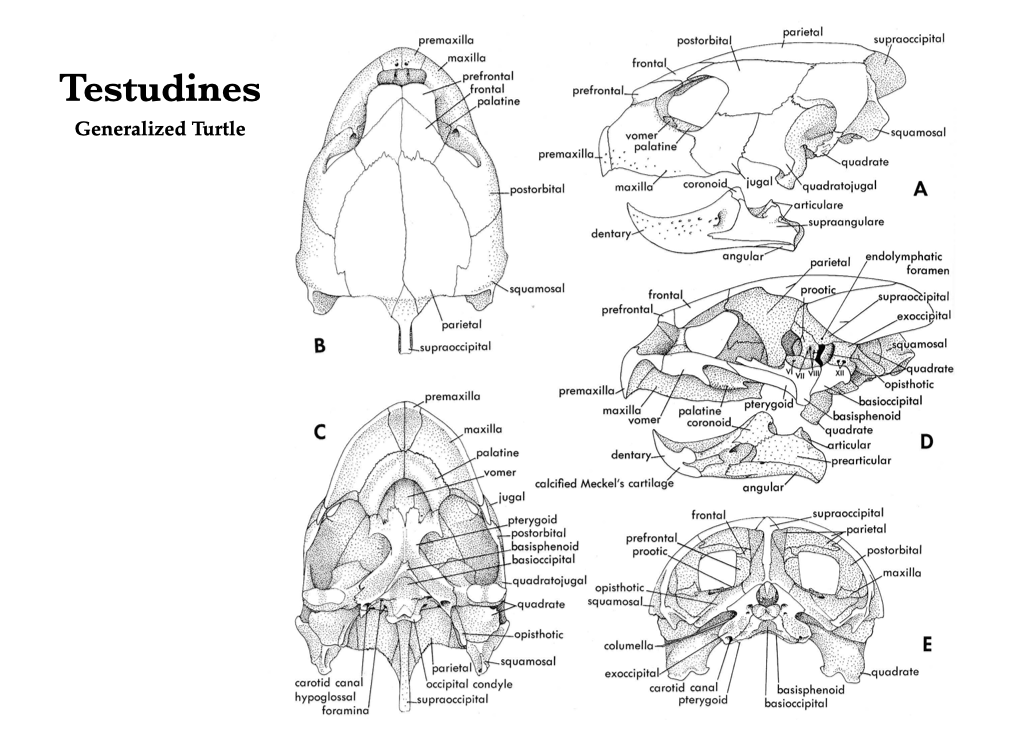
Figure 3.13. Skull of a turtle in several views: lateral (A), dorsal (B), ventral (C), mid-sagittal (D), and caudal (E). Illustrations from Jollie (1962) under CC0 public domain.
Crocodile Skulls (Crocodylia)
Modern crocodylians (crocodiles, alligators, and caimans) have very solid skulls without cranial kinesis. The premaxillae, maxillae, and palatines expand medially, and, with the pterygoids, they form a secondary palate that separates the nasal passageway from the mouth. The choanae are located caudal to the secondary palate. The foramen magnum is encircled by the basioccipital and exoccipitals. The parietals and frontals are fused to their counterparts on the opposite side, forming a single parietal and a single frontal. The nasals overlap the frontals, and the prefrontal contributes to the inner wall of the orbit. Both supratemporal and infratemporal fenestrae are present. The mandible is composed of the dentary, angular, surangular, articular, prearticular, and splenial. A large mandibular fenestra is present, a characteristic of the Archosauria in general.
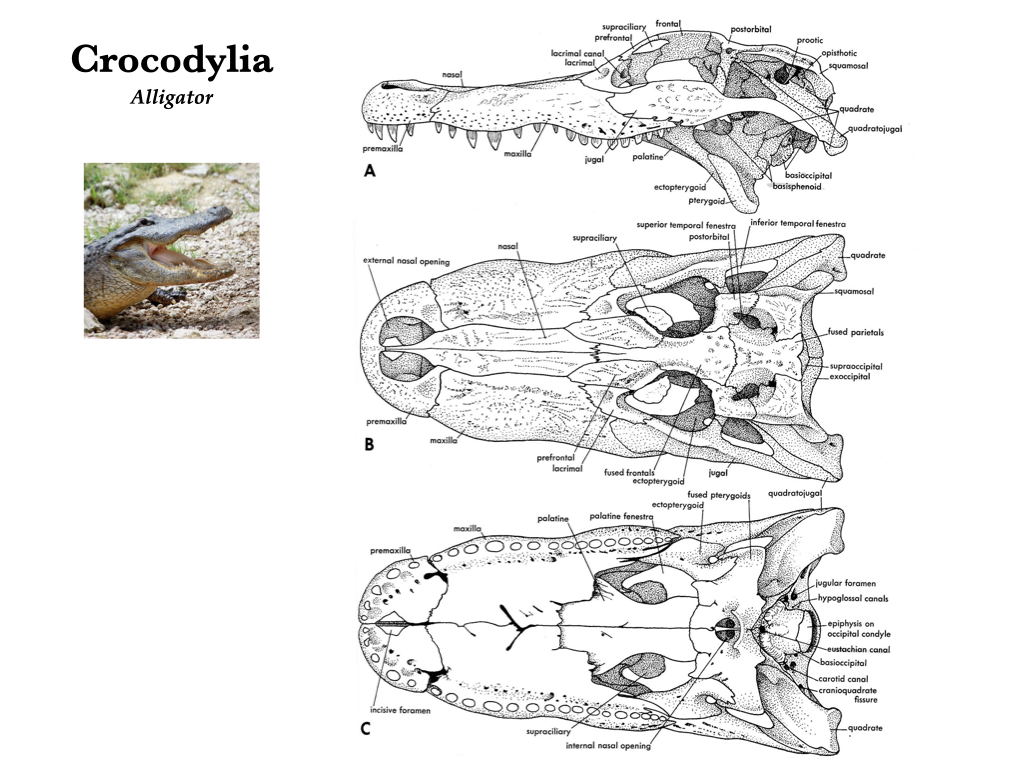
Figure 3.14. Cranium of an alligator (Alligator) in lateral (A), dorsal (B), and ventral (C) views. Illustrations from Jollie (1962) under CC0 public domain; Alligator photo from Flickr user Papyrologist under CC BY-SA 2.0.
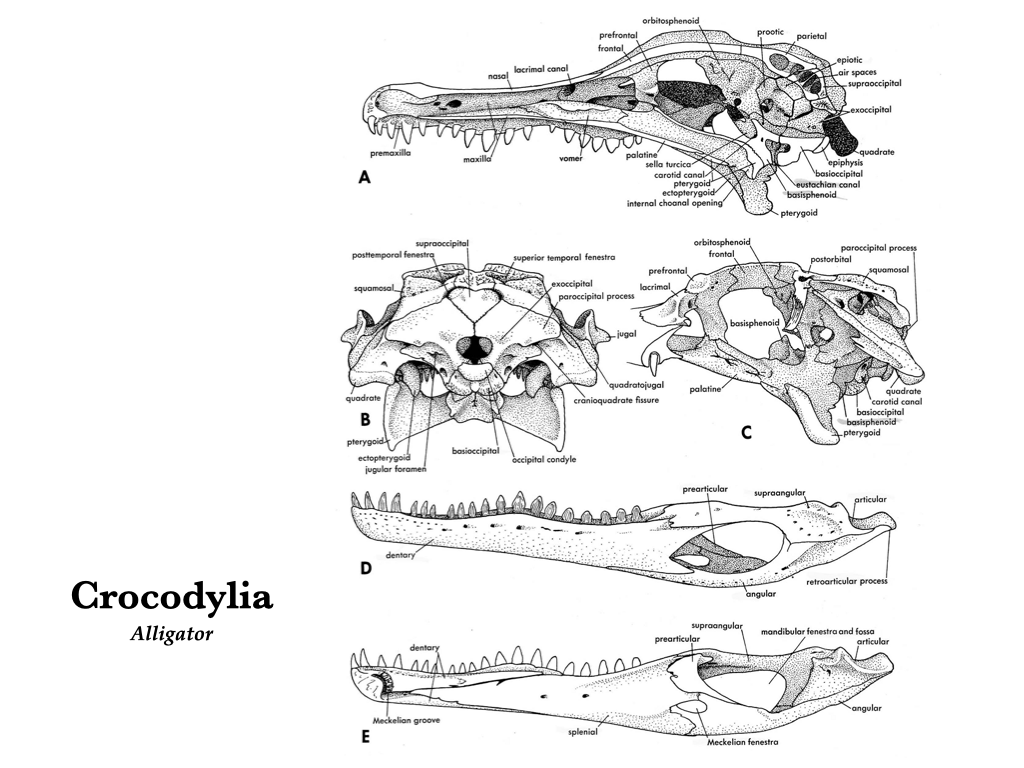
Figure 3.15. More views of an alligator skull: medial (A), posterior (B), lateral with labial arch and quadratojugal removed (C), lateral left mandible (D), and medial right mandible (E). Illustrations from Jollie (1962) under CC0 public domain.
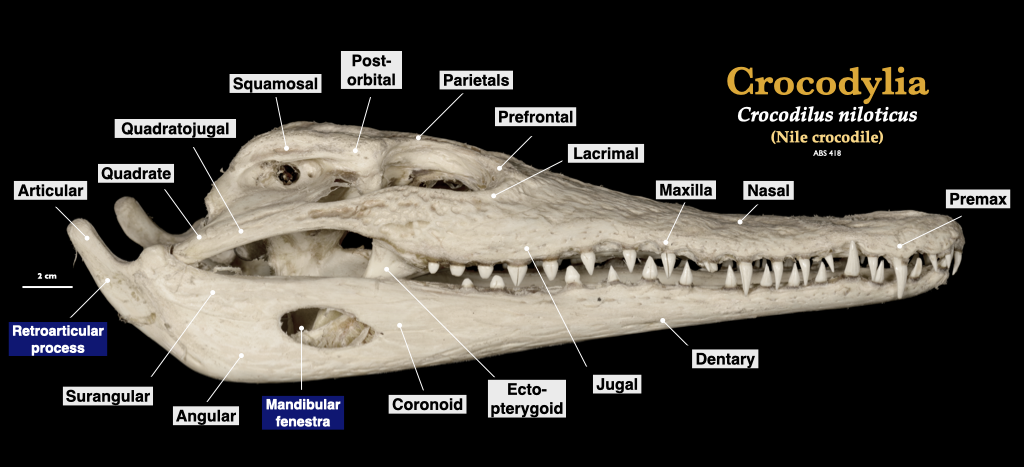
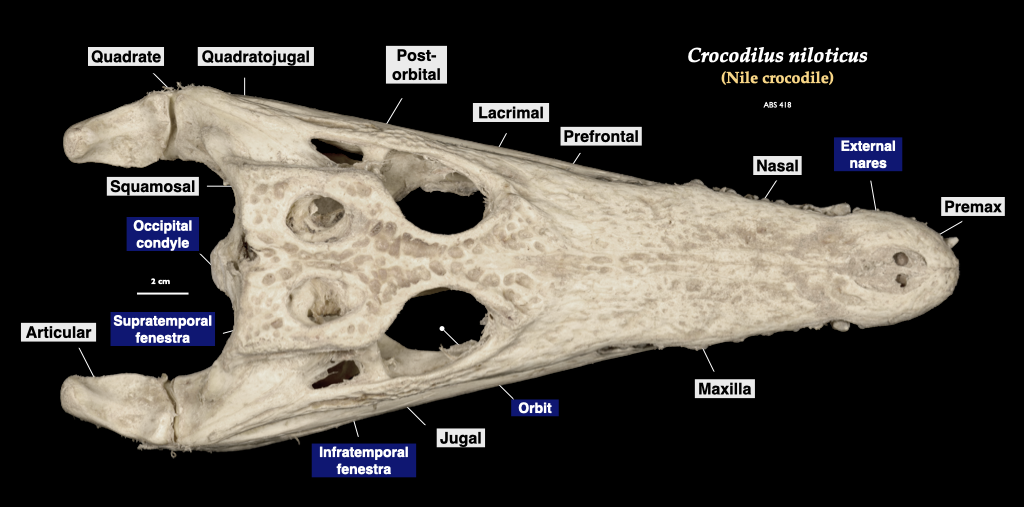
Figure 3.16. Skull of a crocodile (Crocodylus) in lateral and superior view. Crocodylus niloticus specimen from a private collection.
Bird Skulls (Aves)
With the transition to flight, bird skeletons underwent extensive modification. The bones of bird skulls are generally thin, and many are fused, making them rigid. Teeth have been lost in modern taxa and the bones that form the upper and lower jaws are surrounded by a keratinous beak. The mandible is formed from the fusion of the dentary, prearticular, surangular, angular, and splenial, however many of these bones are so tightly fused that the sutures are indistinguishable and only the dentary can be distinguished from the other four. Vision is very important for birds, so the eyes and orbits are large, and many birds have evolved sclerotic bones in the wall of the eyeball. The braincase of birds is expanded relative to other reptiles. A large mandibular fenestra is present in birds as they, too, are archosaurs.
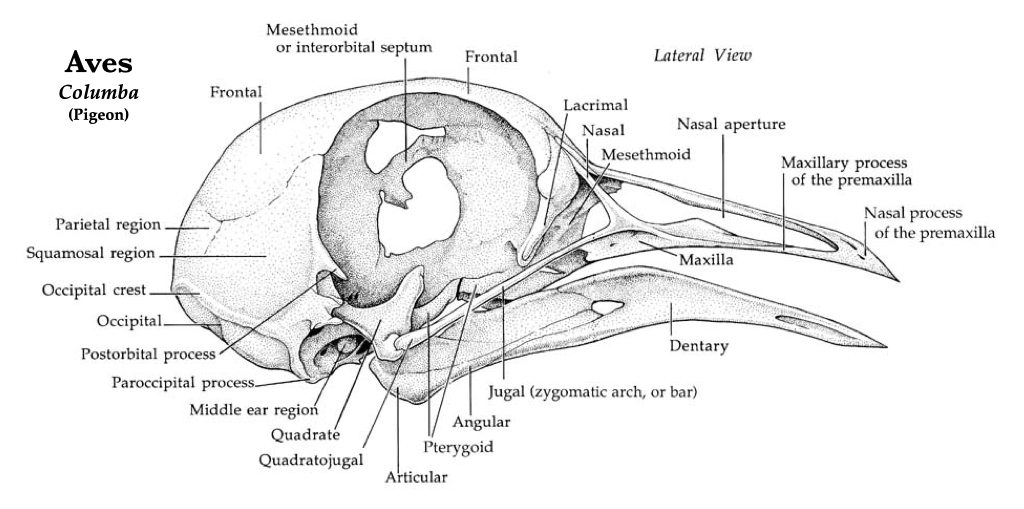
Figure 3.17. Pigeon skull in lateral view. Illustrations by Patrick J. Lynch used with permission of the artist.Figure 3.19. More views of a pigeon skull: dorsal, ventral, posterior (from left to right). Illustrations by Patrick J. Lynch used with permission.
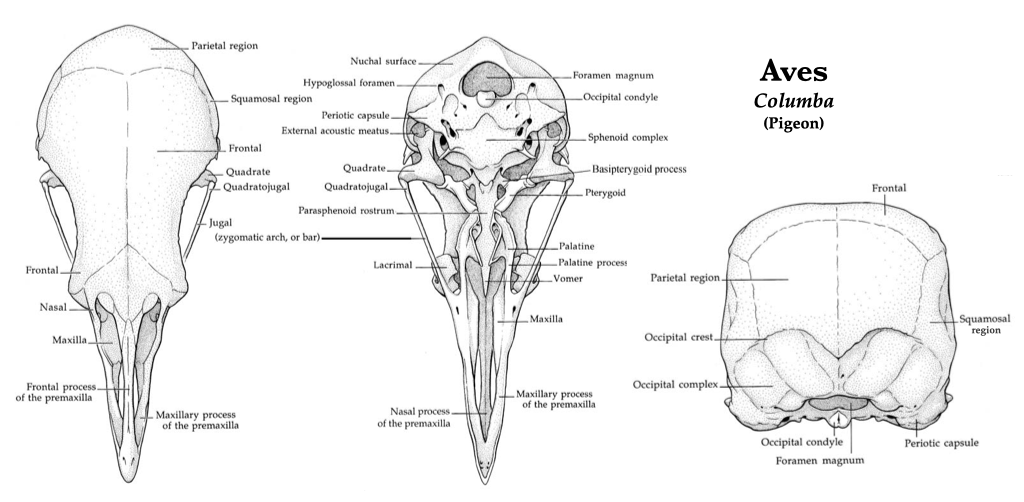
Figure 3.18. Skull of a pigeon (Patagioenas fasciata, MVZ 186085) in lateral view (top) and cranium in inferior view (bottom).
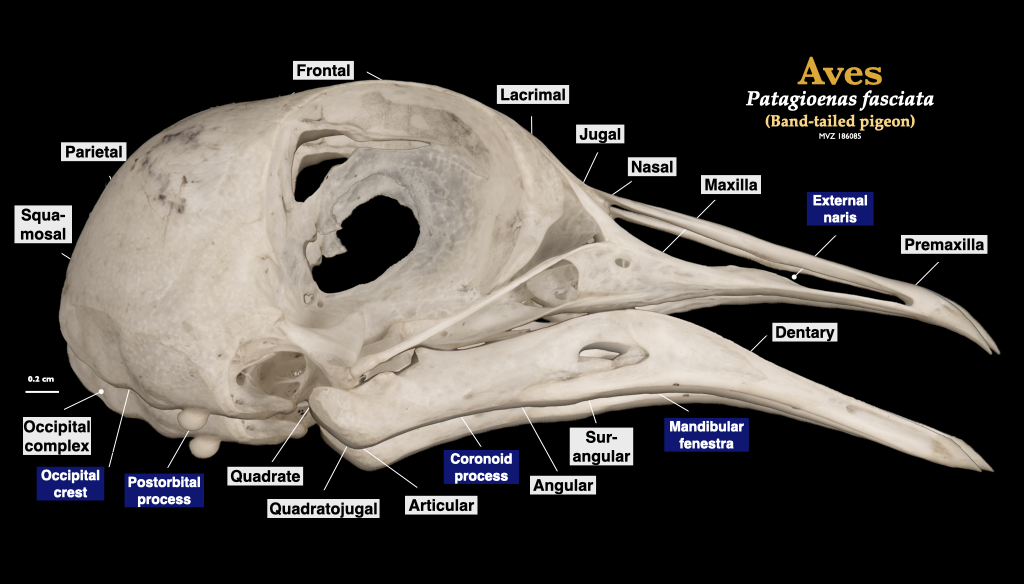
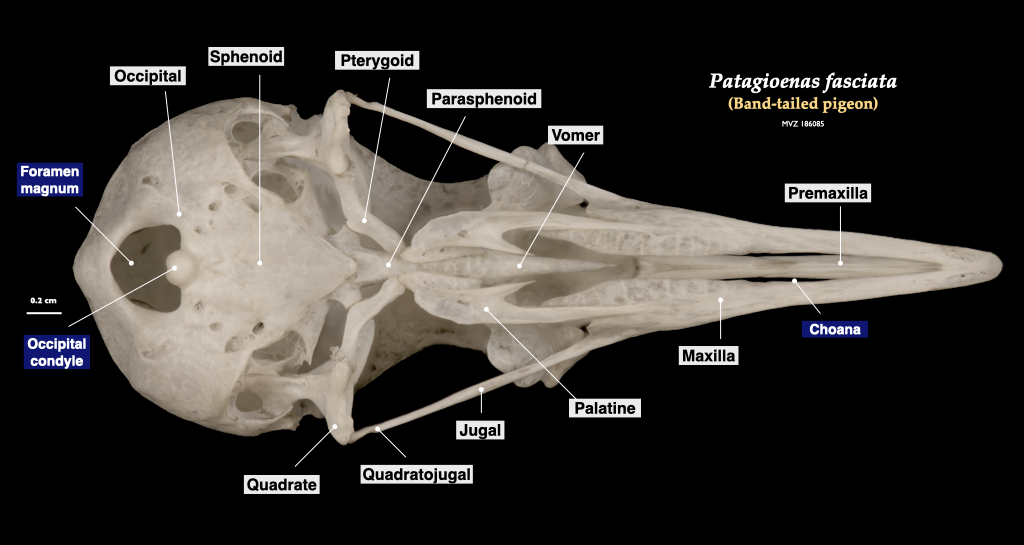
Figure 3.19. Skull of a pigeon (Patagioenas fasciata, MVZ 186085) in lateral view (top) and cranium in inferior view (bottom).








