4 Mammal Skulls
Objectives
- Identify the bones and important features of the mammal skull.
- Distinguish among monotreme, marsupial, and placental mammal skulls based on morphology.
- Explore the evolution of the sense organs, with a focus on the auditory system.
- Study the major morphological differences within the mammalian clades Rodentia and Carnivora.
Overview
In this lab we will focus on the mammal skull. Mammals are endothermic animals with a very high metabolism. Endothermy and high metabolic rates reflect active lifestyles, and mammals are highly active and alert to their surroundings. Many changes to the mammal head are tied to these factors. As a result of their high energetic demands, mammals need to obtain relatively more food than other vertebrates, and they must extract as much energy as possible from this food. Mammals masticate their food efficiently, reducing food items to relatively small particles with high surface area, and this facilitates the action of digestive enzymes and leads to more rapid assimilation. For mastication to be effective, the skull needs to be akinetic, the jaws need to be strong, and the masticatory muscles need to be powerful. Mammals are alert for prey and vigilant against predators, and they have large brains and well-developed sense organs to monitor their surroundings. When you are examining the mammal skulls, notice the robustness of the dentary and the skull overall, and consider the spaces and cavities and foramina that support the neural, visual, olfactory, and auditory systems. After this lab, you will be able to identify the objects listed below in bold face in the articulated skull, and you will learn to identify the objects with asterisks (*) in isolation.
See the terms list here: Terms 4.1
Some notes on mammal skulls
The mammalian skull has several characteristic features:
- Nasal turbinals (aka nasal turbinates, nasal conchae) form from the maxillae, nasals, and/or mesethmoid
- Most circumorbitals are lost, and the postorbital is either reduced or lost altogether
- Two occipital condyles are present
- A secondary palate is present
- The braincase is relatively large
- Three ear ossicles are present and enclosed in an auditory bulla on the basicranium
- Upper teeth are present on the maxilla and premaxilla only
- Mandible composed solely of the dentary
Monotreme skulls can be distinguished from those of other mammals by:
- Absence of auditory bullae
- Absence of teeth in adult individuals (not true for some ancient monotremes)
Marsupial skulls can be distinguished from those of placentals by:
- Participation of the jugal in the jaw joint at the glenoid (mandibular) fossa
- Presence of posterior palatal vacuities
- Auditory bullae derived at least in part from the alisphenoid
- Angular process of the dentary usually inflected medially
- Primitive dental formula of 5134/4134
Placental skulls can be distinguished by:
- Disproportionately large braincase
- Absence of posterior palatal vacuities (there are exceptions)
- Complex auditory bullae without an alisphenoid contribution
- Absence of the jugal in the mandibular fossa
- Primitive dental formula of 3143/3143
Additional Information
Short descriptions of select bones and features
Alisphenoid bones – Wing-like bones in the walls of the temporal fenestrae, caudal to the frontal and orbitosphenoid bones and rostral to the squamosals. In marsupials they are large and contribute to the auditory bullae. An alisphenoid canal, which transmits part of the 5th cranial nerve, may penetrate a bony shelf at the ventral base of the alisphenoid bone. The alisphenoid is homologous to the epipterygoid of teleosts and early amniotes.
Angular process – A process of variable size at the posteroventral edge of the dentary. The angular process is the insertion site for some of the masseter muscles. It is generally large in herbivores, and it is inflected in marsupials and some rodents.
Auditory bullae – Paired bony capsules lateral to the basioccipital and ventral to the squamosal. Each bulla consists of the tympanic bone or a fusion of the tympanic and entotympanic bones. The mastoid and alisphenoid bones may also participate. The tympanic bone supports the ear drum and is homologous to the angular of early amniotes. In eutherians, the tympanic bone is called the ectotympanic bone. The entotympanic bone evolved in mammals and has no homologue in other taxa (it is a neomorph). The bullae protect the middle ear ossicles and facilitate transmission of sound to the inner ear.
Condyloid process – The projection on the posterodorsal portion of the dentary that bears the mandibular condyle, which articulates with the squamosal at the glenoid fossa.
Coronoid process – The projection extending dorsally from the dentary into the temporal fossa. The coronoid process provides surface area for muscle attachment (it is the insertion site for the temporalis), and it is large in mammals that require a strong bite force, such as carnivorans, and it is small or absent in some herbivores.
Frontal bones – Paired bones posterior to the nasals and dorsal to the maxillae. In the odontocetes, the frontal bones are telescoped posteriorly. The antlers and horns of artiodactyls grow from the frontal bones.
Jugal bones – The jugals form the anterior portion of the zygomatic arch. In marsupials, they participate in the mandibular fossa. In rodents, they contact the lacrimals. When the zygomatic arch is incomplete, it is usually through the reduction or loss of the jugal (this occurs, for example, in some monotremes, in shrews, and in pangolins).
Lacrimal bones – Each lacrimal is located on or adjacent to the anterior base of the zygomatic arch at the dorsal edge, and each is perforated by a lacrimal foramen with a tear duct. Lacrimal bones are absent in monotremes and pangolins.
Masseteric fossa – The depression on the lateral surface of the dentary, ventral to the coronoid process. The bulk of the masseter muscle inserts here.
Mastoid bones – Small, usually obscure bones just lateral to the caudal margin of the auditory bullae. The mastoid is a part of the periotic, a general term for the complex of bones around the ear, including the petrosal and mastoid (also called the petromastoid). In some mammals it is reduced to a process (mastoid process), in heteromyid rodents, the mastoid forms part of the inflated bulla.
Maxillae – Tooth-bearing bones that make up a large part of the rostrum and palate. The maxillae are especially large in cetaceans.
Nasal bones – Paired bones forming the rostral roof of the nasal cavities. Reduced in odontocetes (toothed whales), absent in some sirenians (dugongs).
Occipital condyles – Paired swellings of the exoccipital bones adjacent to the foramen magnum, each of which articulates with the atlas.
Parietal bones – Paired bones posterior to the frontals. Enlarged in anthropoids.
Periotic (or petromastoid) – A bone that is the result of a fusion of the prootic, epiotic, and opisthotic.
Postparietal (Interparietal) bone – A single, often triangular bone of the braincase posterior to the interparietal suture. Often fused with the occiput in adult individuals, but distinct in many rodents.
Premaxillae – Paired, tooth-bearing bones that form the rostral roof of the mouth and the ventral portion of the external nares. The upper incisors always reside in these bones. Absent in some bats. Fused and difficult to observe in humans.
Squamosal bones – Paired bones located lateral and ventral to the parietal bones. The squamosals contribute to the jaw joint in mammals. The squamosal may be called the temporal bone when it is fused with the periotic; fusion of the squamosal with the periotic occurs in most mammals except marsupials and cetaceans. In humans, the temporal bone also includes the tympanic, and it completely encloses the ear.
Temporal fenestra – The opening posterior to the orbits that is bordered laterally by the zygomatic arches. In most synapsid mammals, the temporal fenestra and orbit are connected. In some taxa, a temporal bar may encircle the orbit; in some primates and in horses, the temporal fenestra and the orbit are separated by a postorbital plate.
Tympanic bone – See the information above on the auditory bullae.
Zygomatic arches – Arches on the sides of the cranium that form the lateral and ventral borders of the orbits and temporal fenestrae. Several bones may contribute to the formation of the zygomatic arch (or zygoma), particularly the maxilla, jugal, and squamosal. In mammals with reduced masseteric musculature, the arches may be incomplete or absent.
Mammalian hearing apparatus
In mammals, the bones of the ancient splanchnocranium (articular, quadrate, and hyomandibula) are modified into middle ear ossicles (malleus, incus, and stapes, respectively), and these bones form a sophisticated system to amplify sound vibrations received through air. The inner ear of therian mammals is housed in an auditory bulla (monotremes do not have auditory bullae). In metatherians, the alisphenoid bones participate to some degree in the bullae. In eutherians, the bullae are formed by the ectotympanic and the entotympanic, without participation of the alisphenoid. Make sure that you can trace the evolution of the mammalian hearing apparatus from its ancient origin.
Monotremata
Note that the skull of the young echidna (below) is not entirely fused because the specimen depicted is young. As in the other extant monotremes, the auditory bullae and teeth are absent, and the maxillae and palatines are elongate.
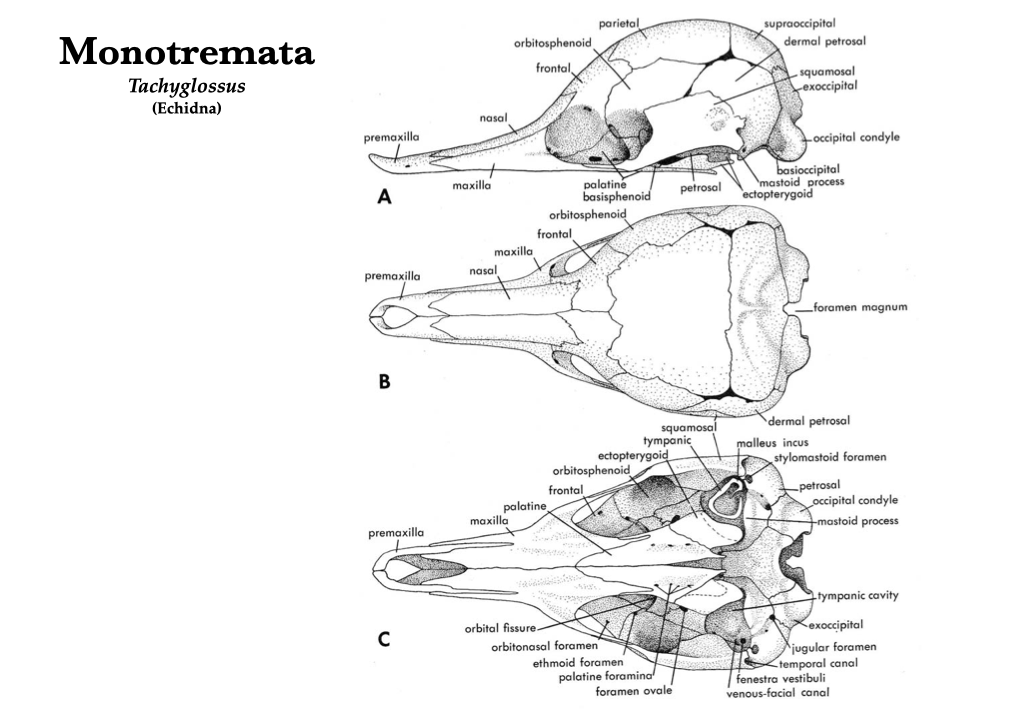
Figure 4.1. Skull of a young echidna (Tachyglossus aculeatus): lateral (A), dorsal (B), and ventral (C) views. Illustrations from Jollie (1962) under CC0 public domain.
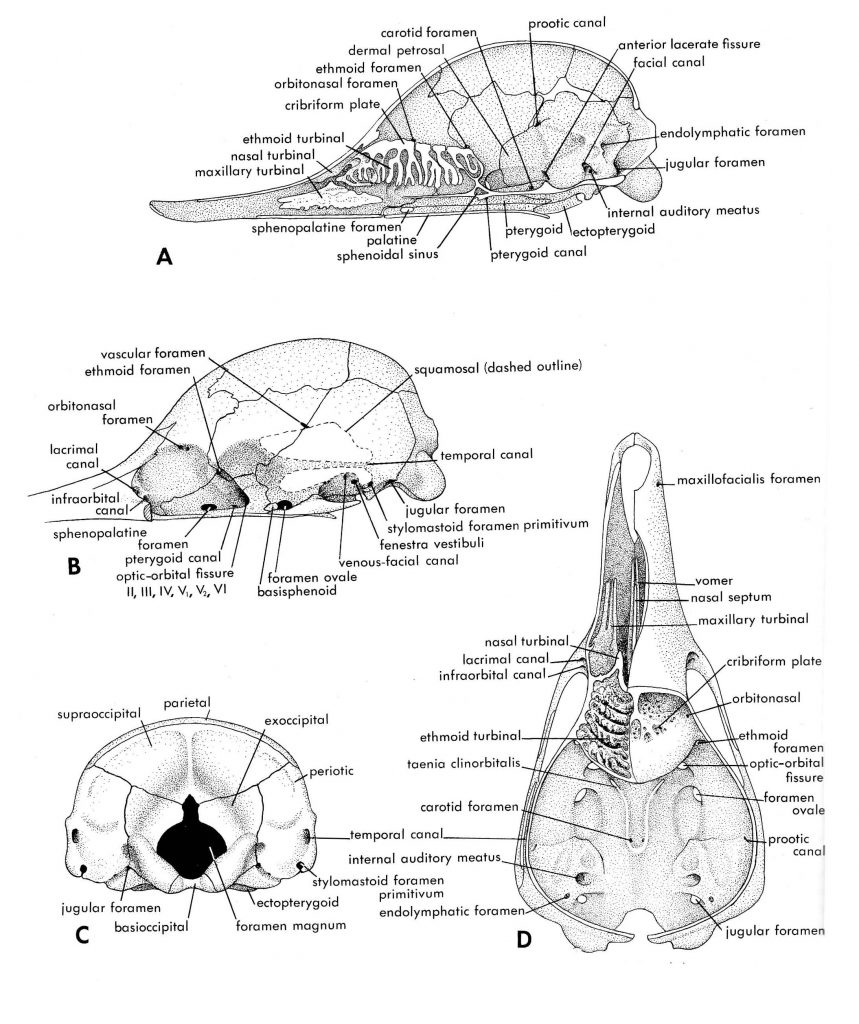
Figure 4.2. More views of a young echidna: medial (A), lateral with zygoma and squamosal removed (B), posterior (C), dorsal with cranial roof removed (D). Illustrations from Jollie (1962) under CC0 public domain.
Marsupialia
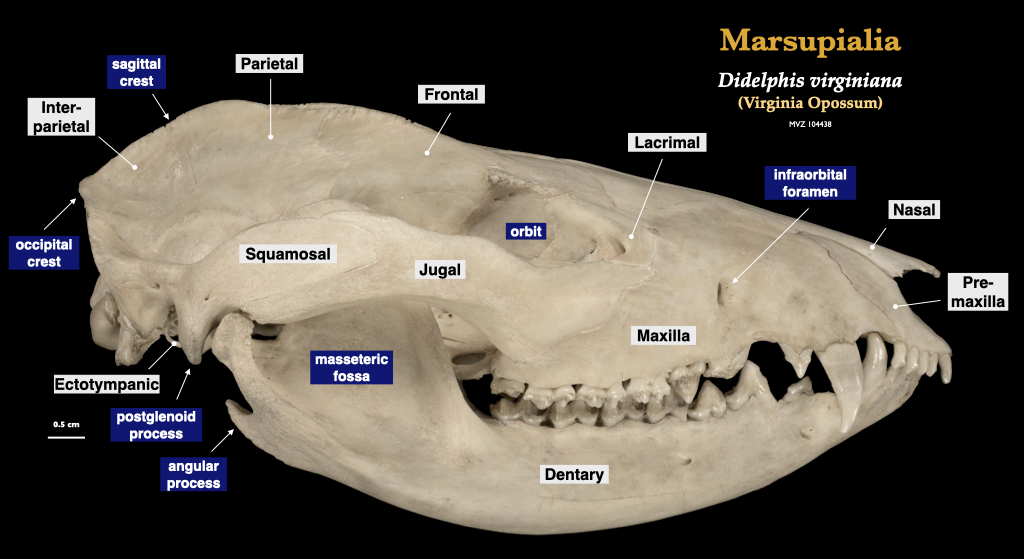
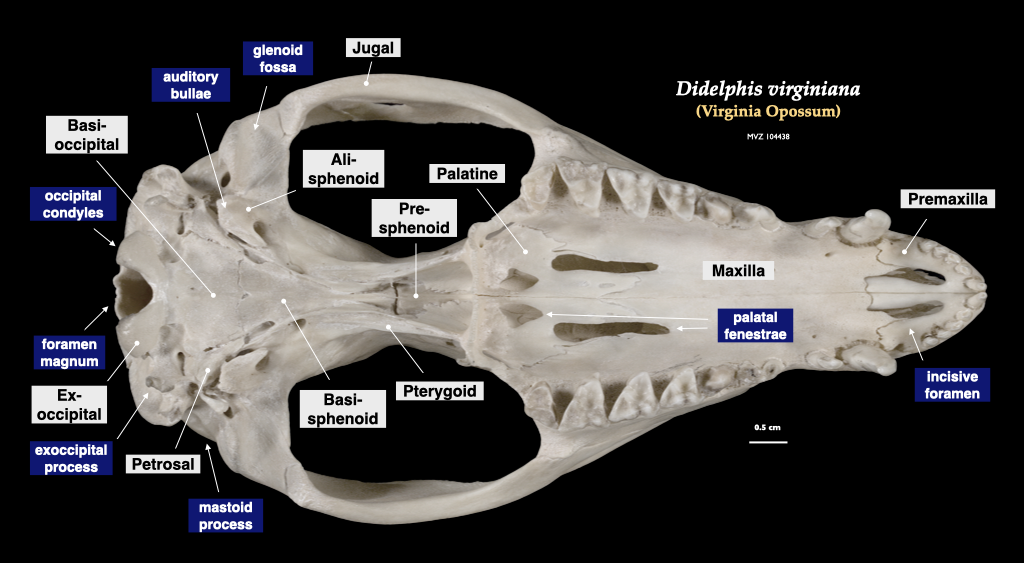
Figure 4.3. Skull of a common opossum (Didelphis virginiana specimen, MVZ 104438) in lateral view, cranium in inferior view.
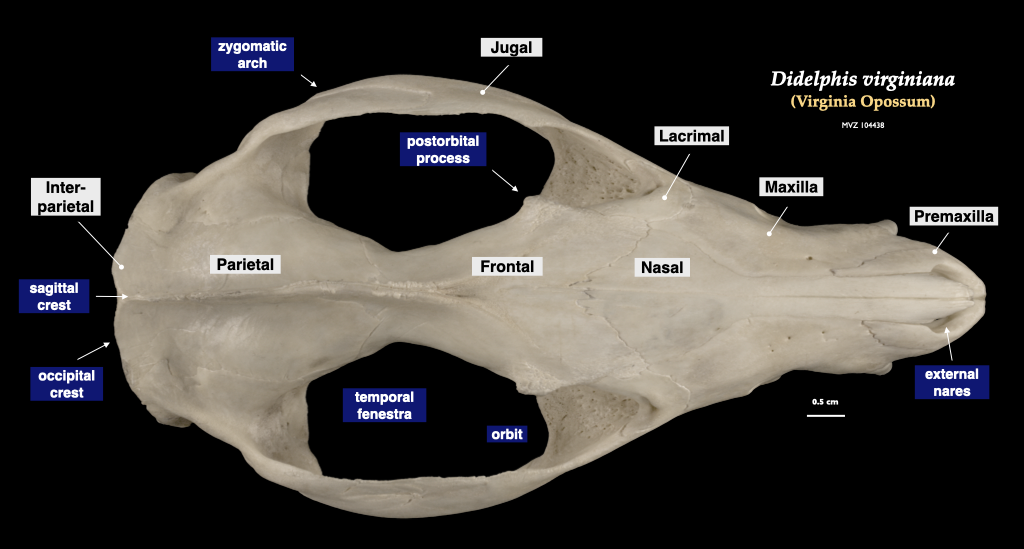
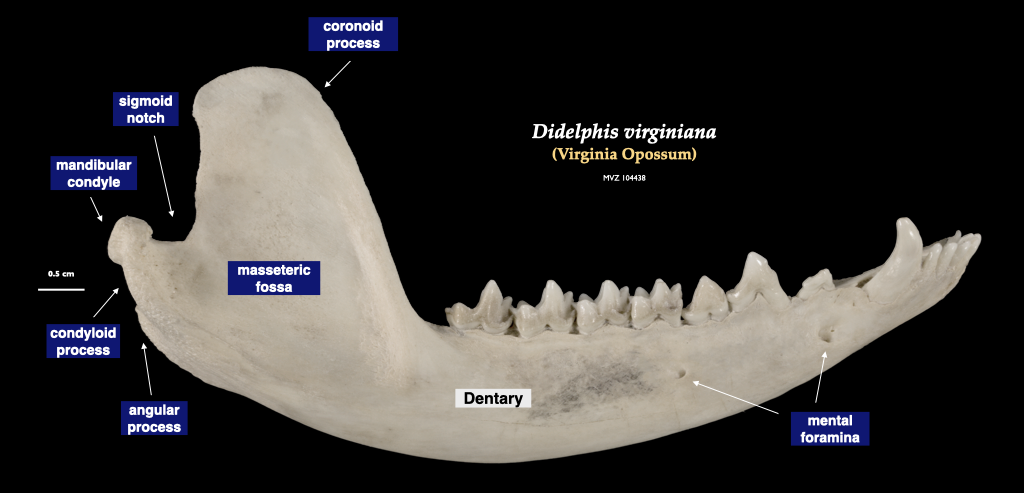
Figure 4.4. Cranium of an opossum in superior view, mandible in lateral view. Didelphis virginiana specimen, MVZ 104438.
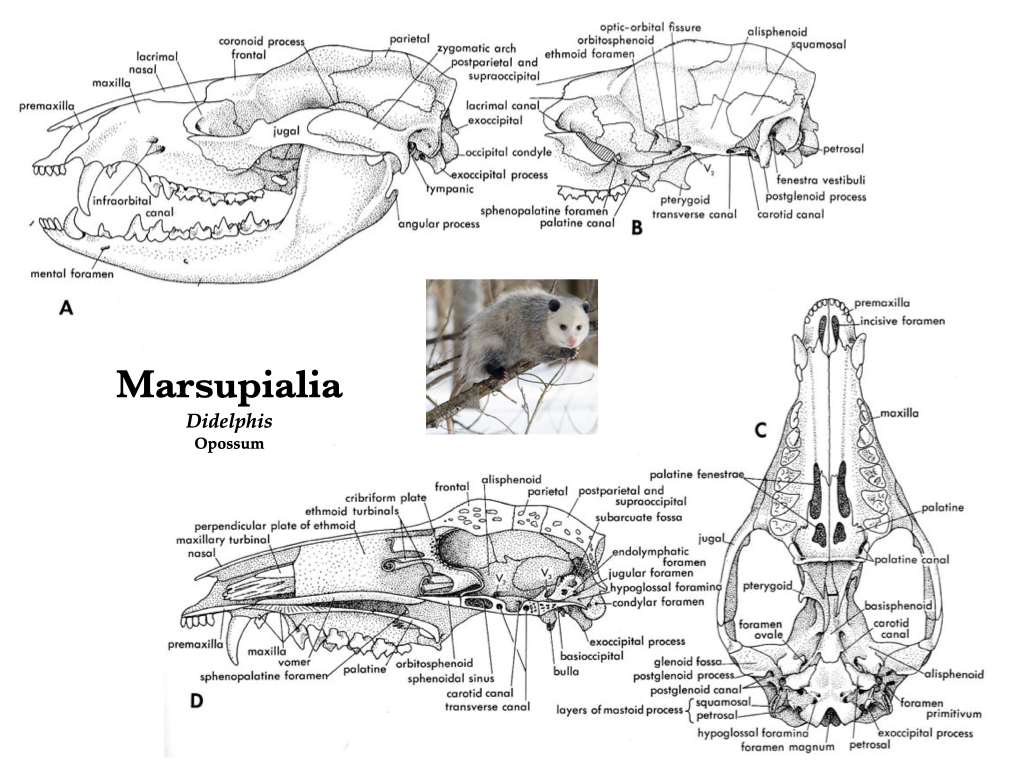
Figure 4.5. Skull of the common opossum (Didelphis virginiana) in several views: lateral (A); lateral with zygomatic arch removed (B), ventral (C), and mid-sagittal (D). Illustrations from Jollie (1962) under CC0 public domain; opossum photo from Wikipedia user Cody Pope under CC BY-SA 2.5.
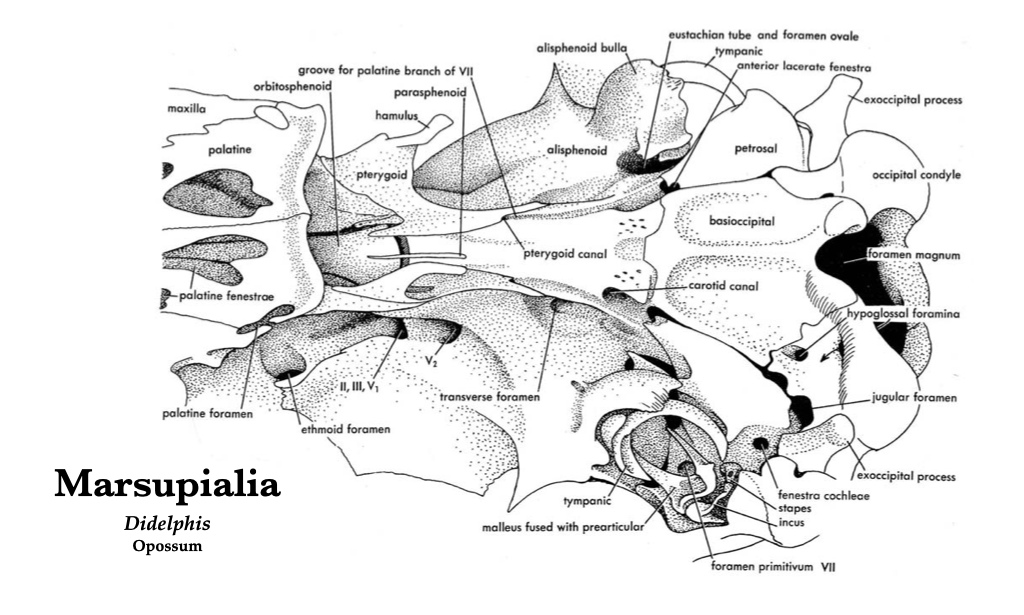
Figure 4.6. Posteroventral view of the opossum skull. Illustration from Jollie (1962) under CC0 public domain.
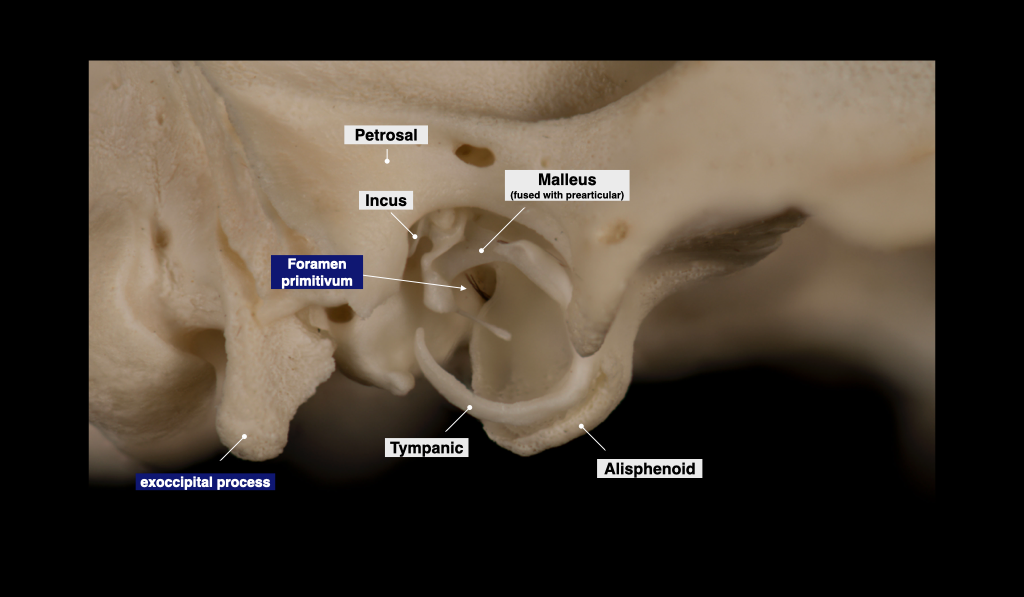
Figure 4.7. Tympanic region of the opossum skull. Didelphis virginiana specimen from a private collection.
Placentalia
The domestic cat skull is a convenient reference point for the morphology of the Eutheria, particularly because these skulls are available in many teaching collections. Compare the cat skull to the skull of a dog, tracking homologous structures. Contrast the overall shape of the cat and dog skulls (e.g., short rostrum versus long rostrum), and the adaptations of the cat dentition to hypercarnivory.
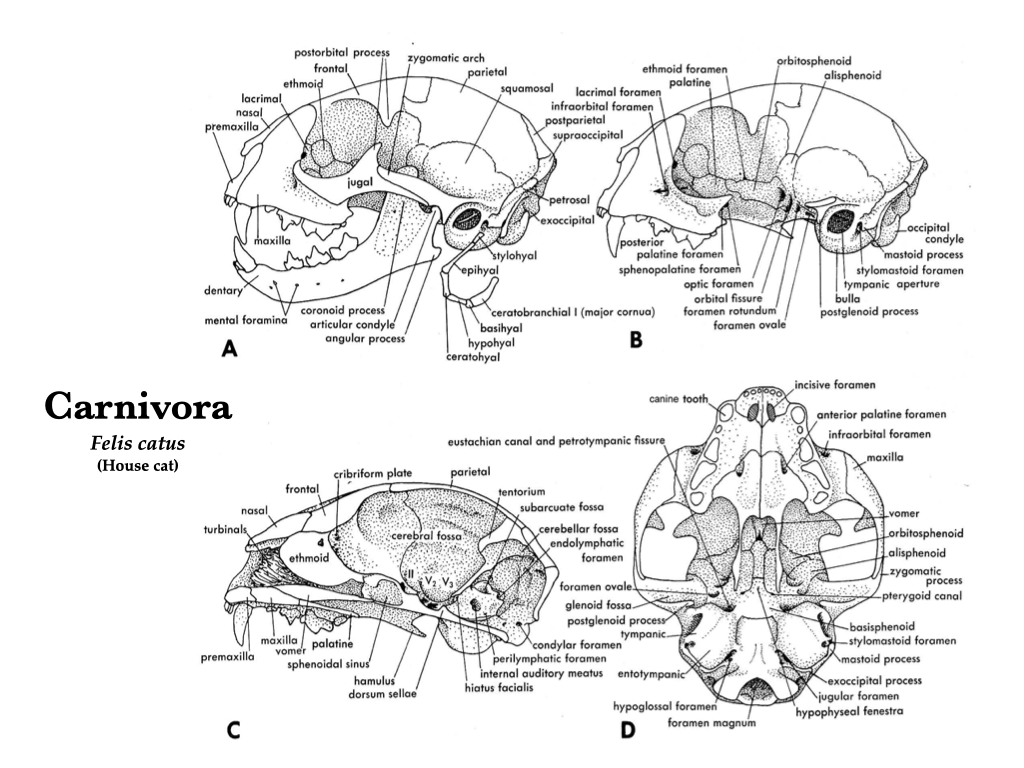
Figure 4.8. Skull of a domestic cat (Felis catus) in several views: lateral (A), lateral with zygomatic arch removed (B), mid-sagittal (C), and ventral (D). Illustrations from Jollie (1962) under CC0 public domain.
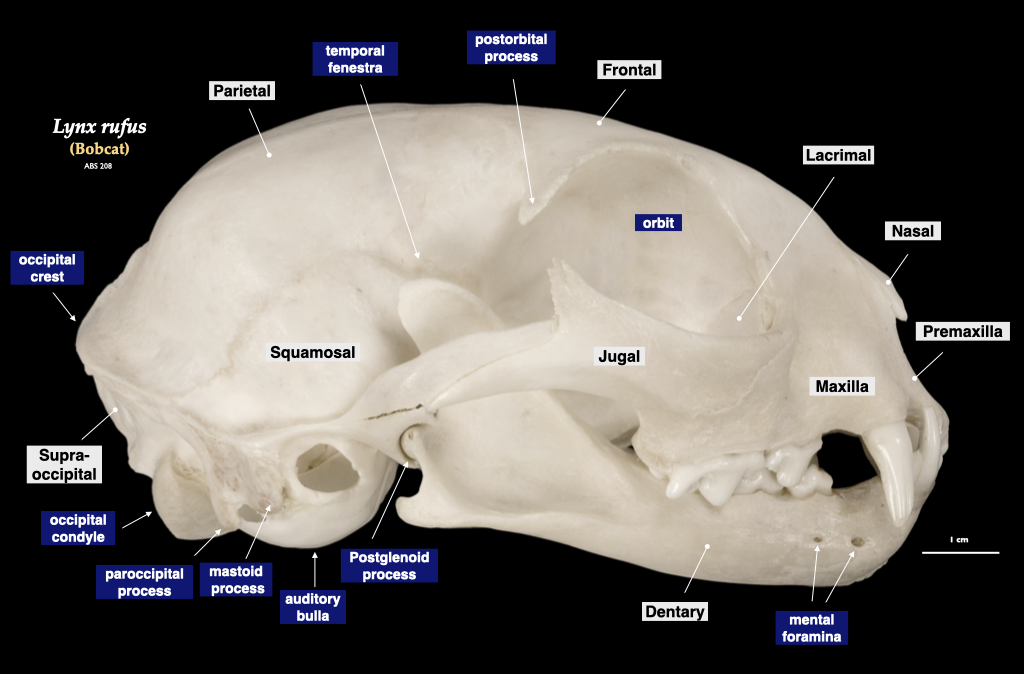
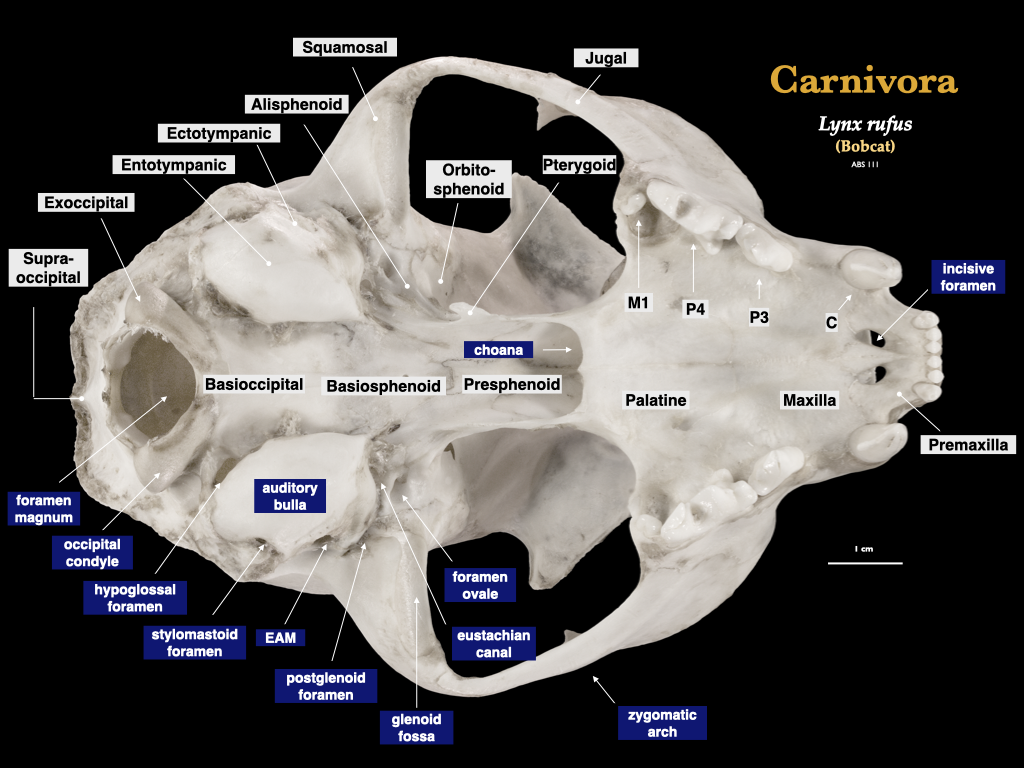
Figure 4.9. Skull of a bobcat (Lynx rufus) in lateral view (top), cranium in inferior view (bottom). Specimen from a private collection.
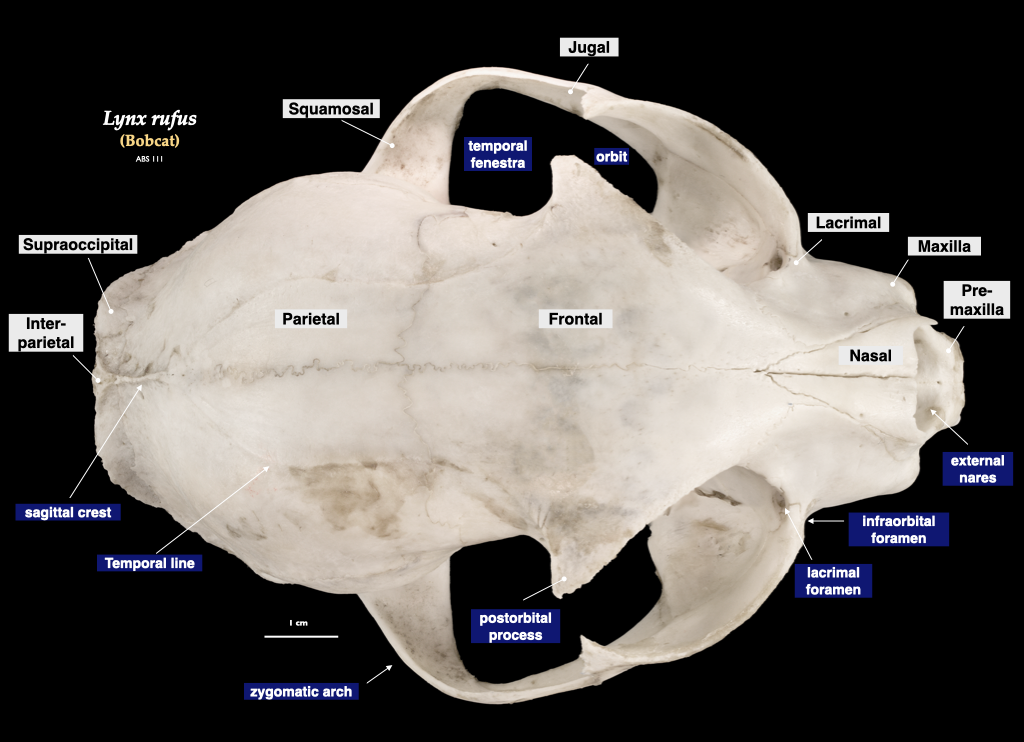
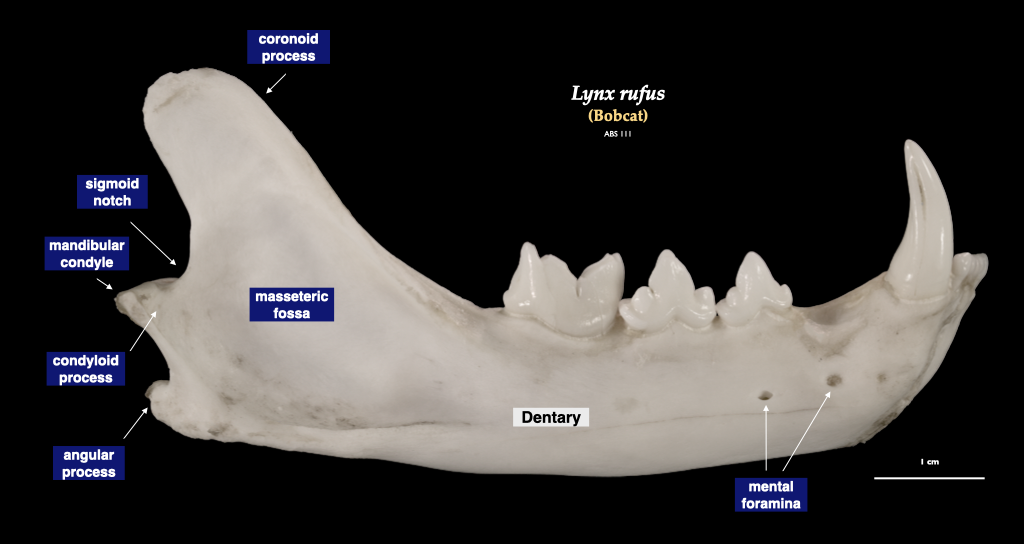
Figure 4.10. Cranium of a bobcat (Lynx rufus) in superior view (top), right mandible in buccal view (bottom). Specimen from a private collection.
Rodent skull and jaw morphology
Rodents have evolved unique jaw muscle configurations, probably related to the unusual way they use their gnawing incisor teeth. There are four major categories of rodent skull configurations related to the masseter muscles, and two major types of mandibular morphology.
Muscle configurations
- Protrogomorphy is a configuration where the masseter muscle originates exclusively on the zygomatic arch, which generally leads to a transverse chewing motion. This is the primitive condition that is present in most non-rodent mammals and in the mountain behavior (Aplodontia).
- Sciuromorphy is a configuration where most of the masseter muscle originates on the zygomatic arch, but a branch of the lateral masseter attaches to the lateral surface of the rostrum. The superficial masseter also originates on the rostrum. This facilitates a rostrocaudal jaw motion, useful both in gnawing with the incisors and grinding with the cheek teeth.
- Hystricomorphy is where the medial masseter originates on the lateral surface of the rostrum and runs medial to the zygomatic arch. In hystricomorphs, the medial masseter passes through a greatly enlarged infraorbital foramen.
- Myomorphy is the condition in which the medial masseter originates on the lateral surface of the rostrum and passes through the infraorbital foramen (as in hystricomorphs) and an extension of the lateral masseter originates on a zygomatic plate at the anterior end of the zygomatic arch. Myomorphy has evolved multiple times in rodents.
Mandibular morphology
- Sciurognathy is where the plane of the angular process is aligned with the plane of the dentary, and it is the primitive condition.
- Hystricognathy is where the plane of the angular process is not aligned with the plane of the dentary, and it is the derived condition.
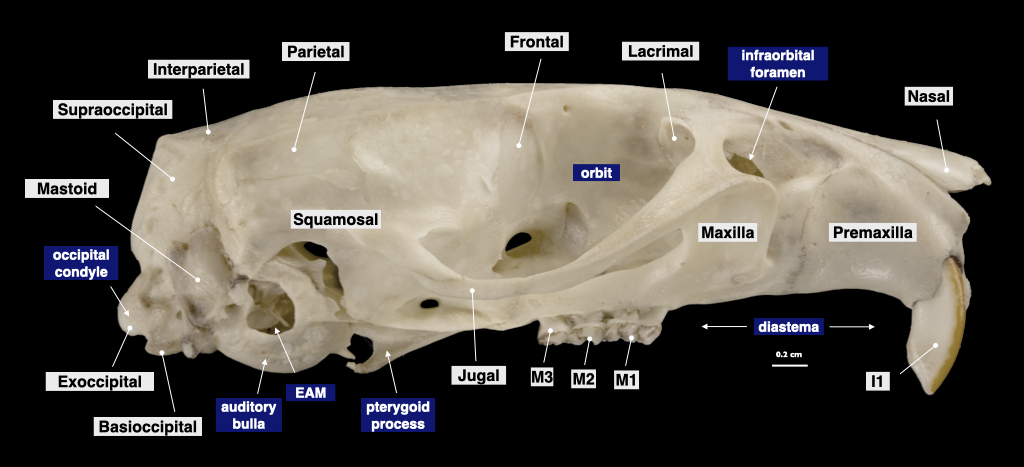
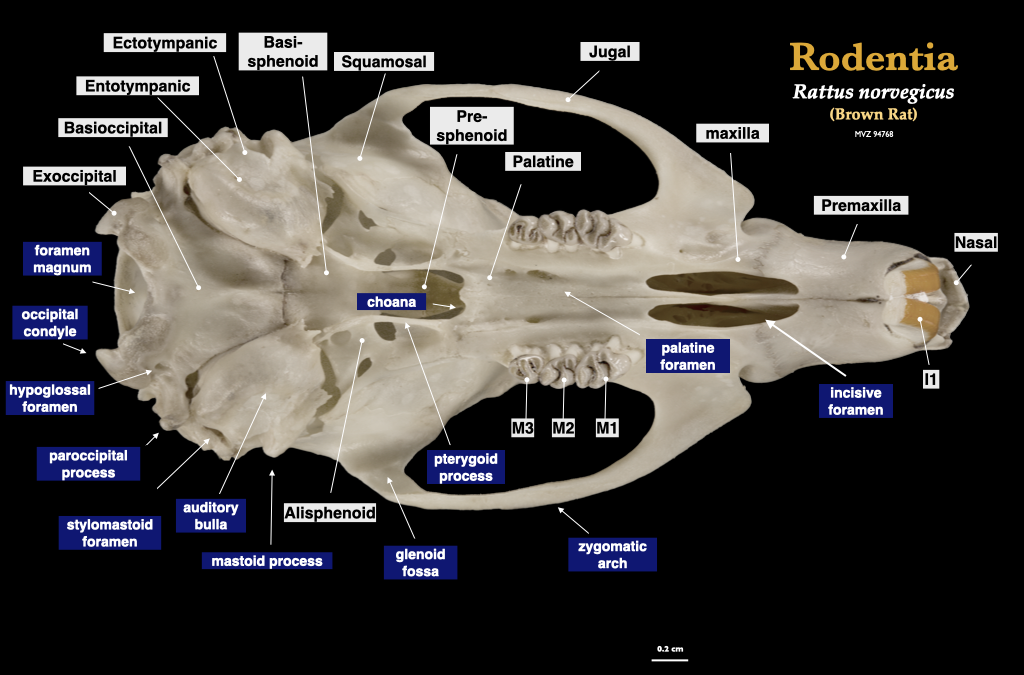
Figure 4.11. Cranium of a rat (Rattus norvegicus, MVZ 94768) in lateral (top) and inferior (bottom) views.
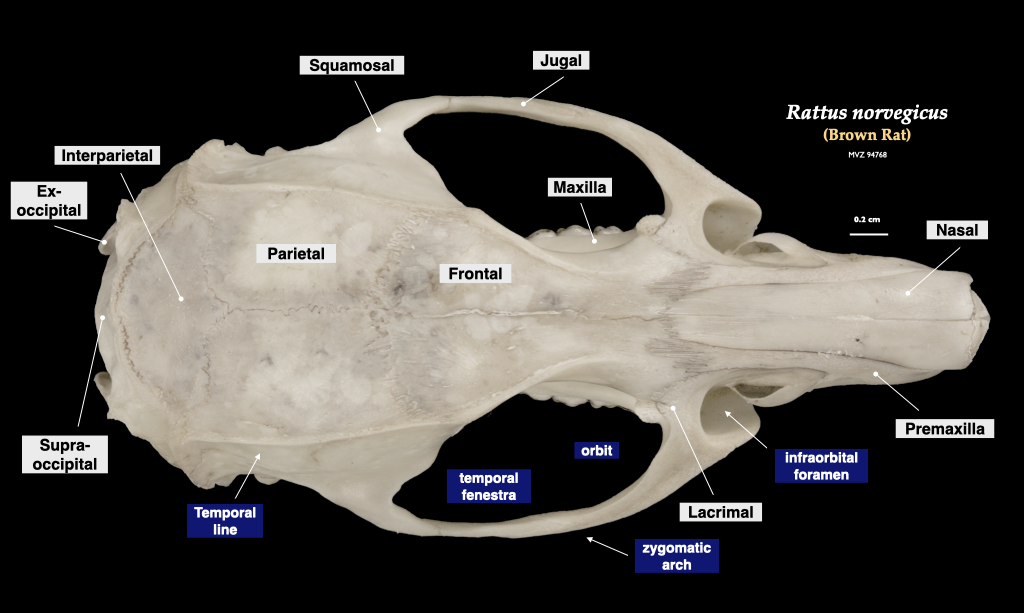
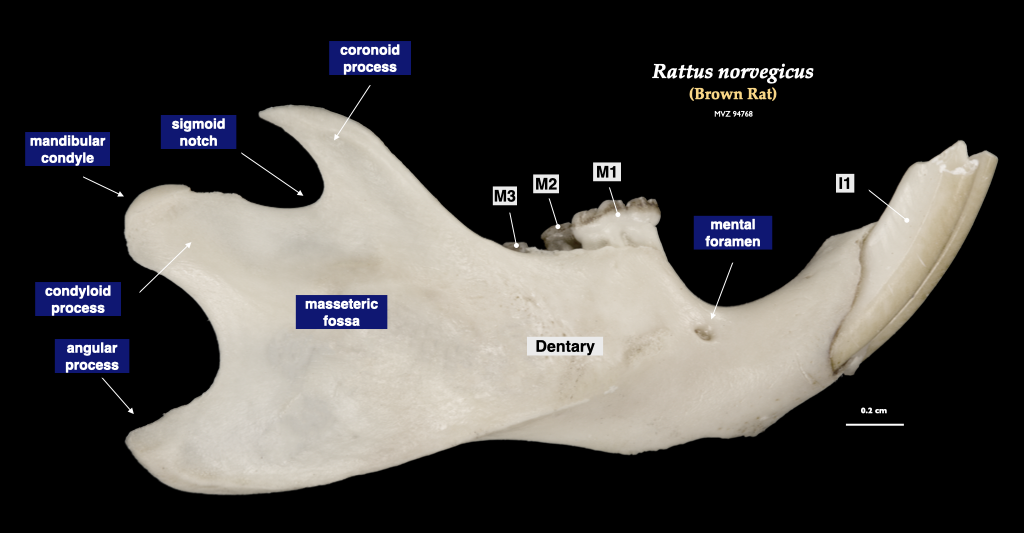
Figure 4.12. Cranium of a rat (Rattus norvegicus, MVZ 94768) in superior view (top) and right mandible in lateral view (bottom).
Cetacean skulls
Porpoises, dolphins, and whales possess skulls that are radically modified for an aquatic existence. The face is “telescoped” with an elongate rostrum and external nares that sit atop the cranium.
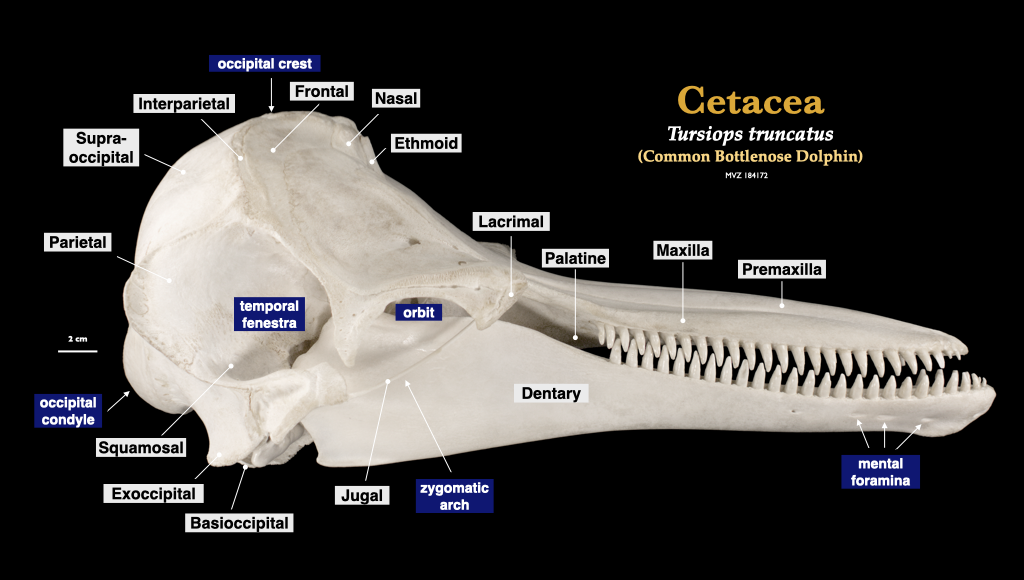
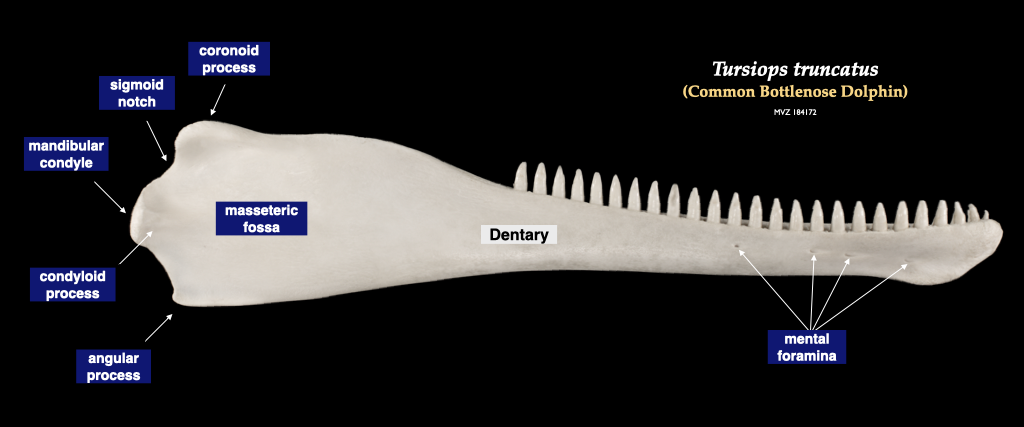
Figure 4.13. Skull of a common dolphin (Tursiops truncatus, MVZ 184172) in lateral view (top), right mandible in buccal view.
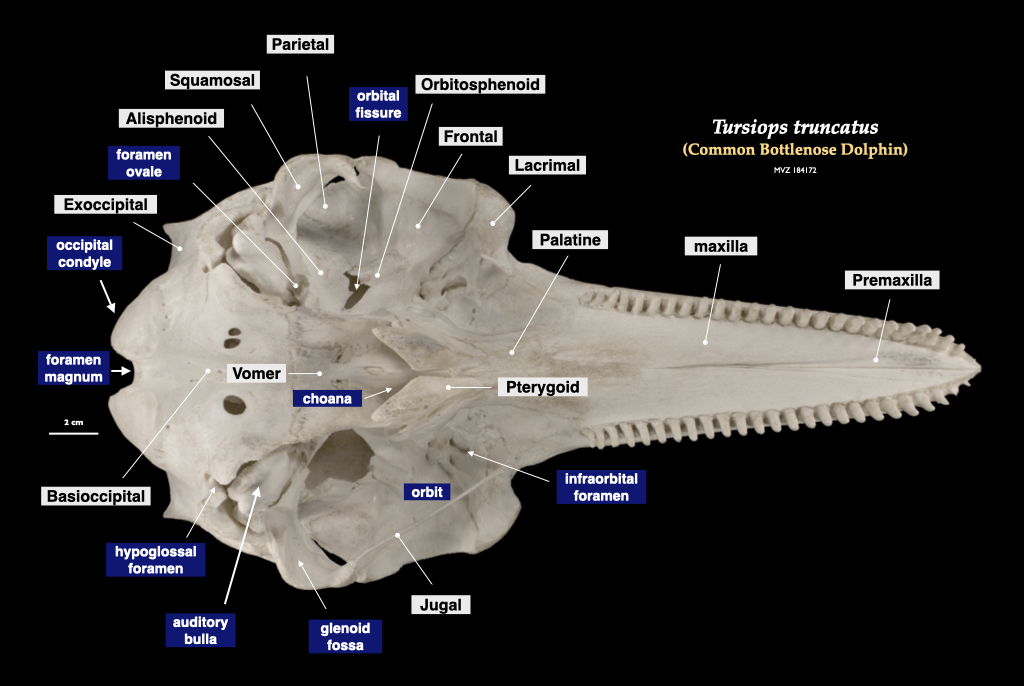
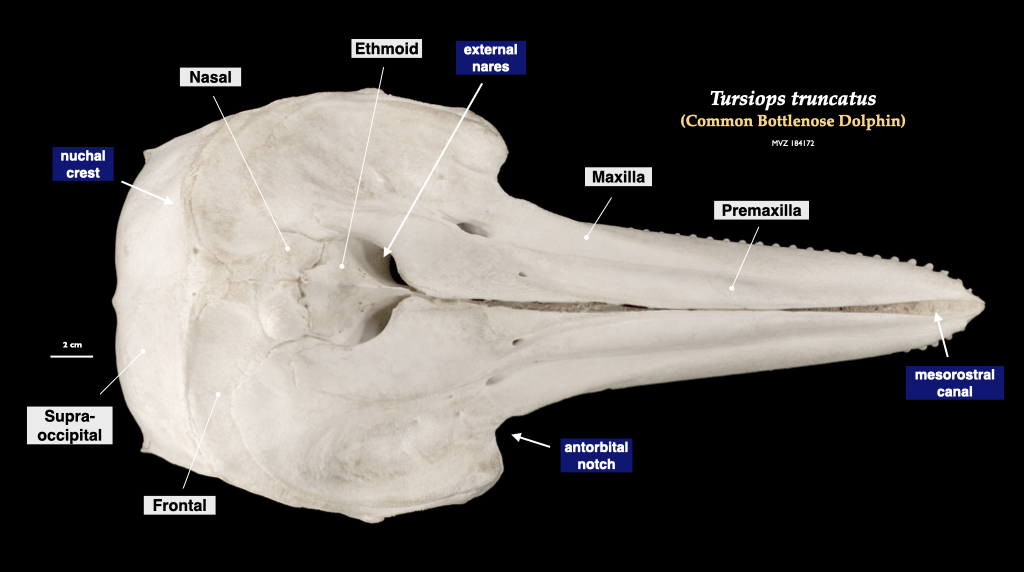
Figure 4.14. Cranium of a common dolphin (Tursiops truncatus, MVZ 184172) in inferior view (top) and superior view (bottom).
Human skull morphology
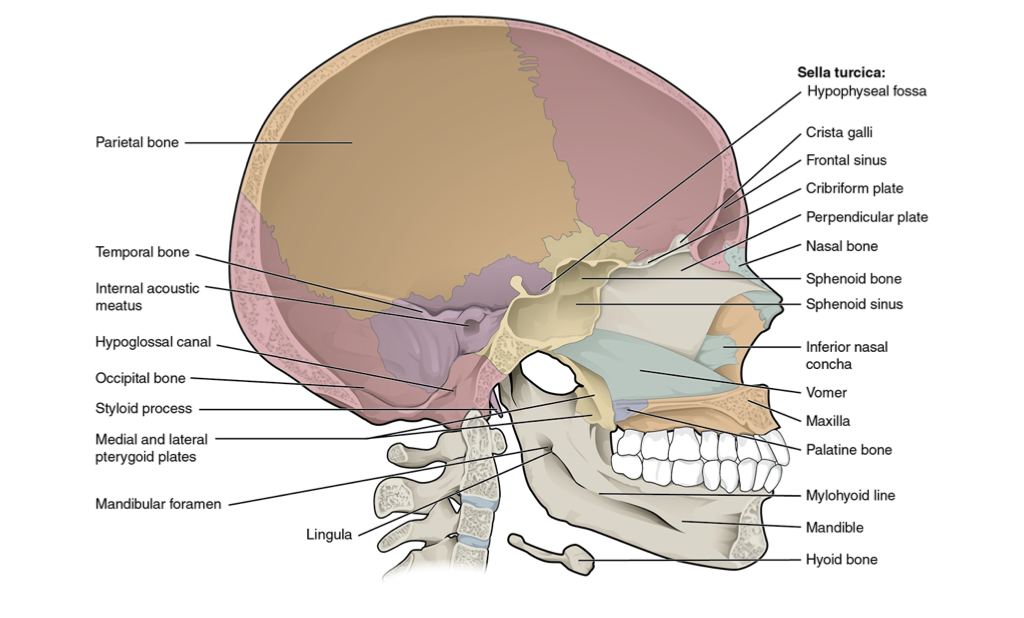
Figure 4.12. Sagittal section of skull from Wikipedia user OpenStax College under CC BY 3.0.
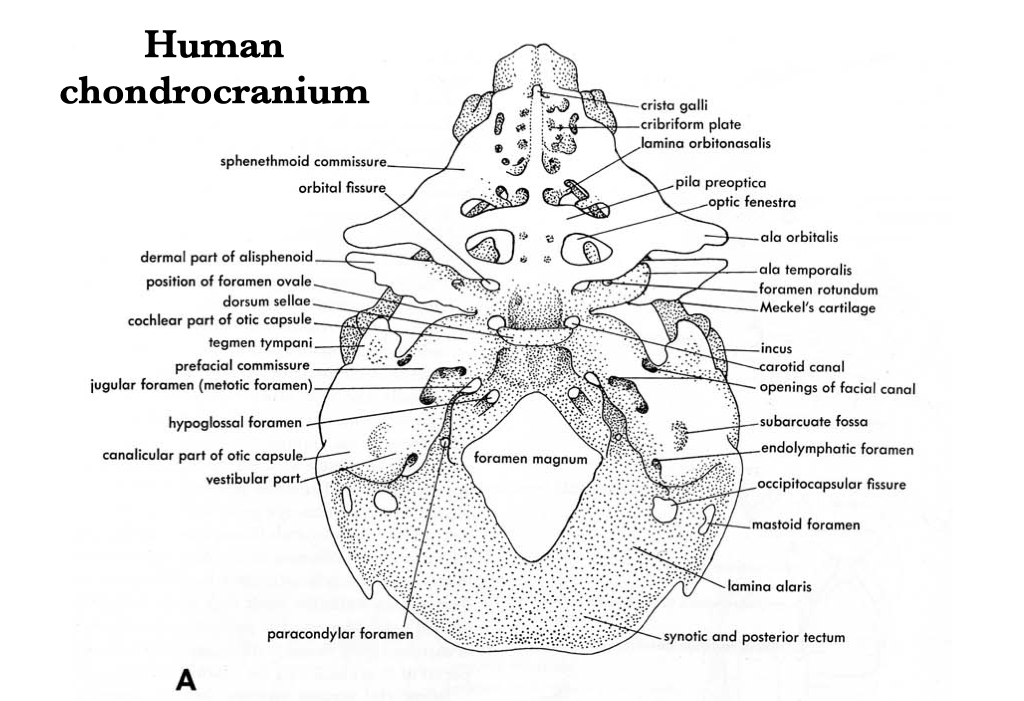
Figure 4.13. Illustration from Jollie (1962) under CC0 public domain.
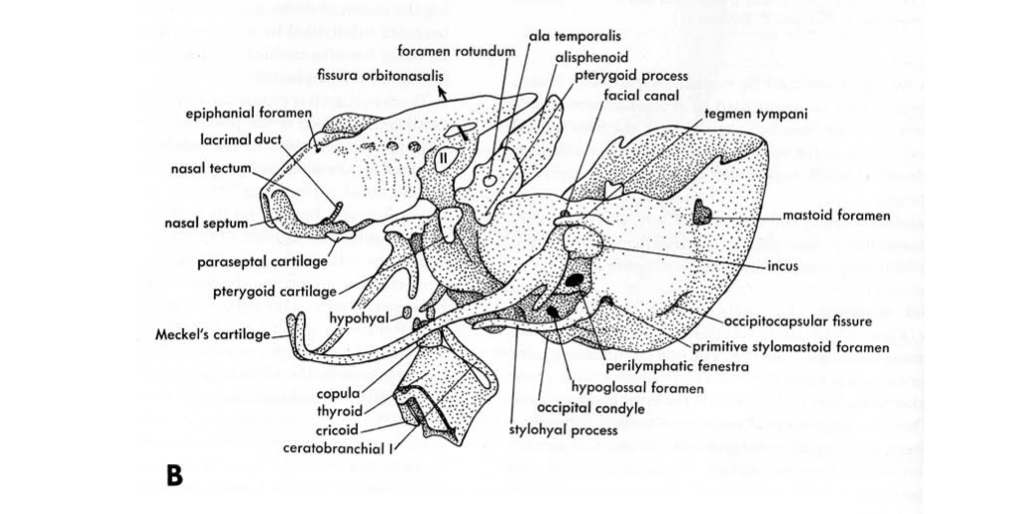
Figure 4.14. Illustration from Jollie (1962) under CC0 public domain.
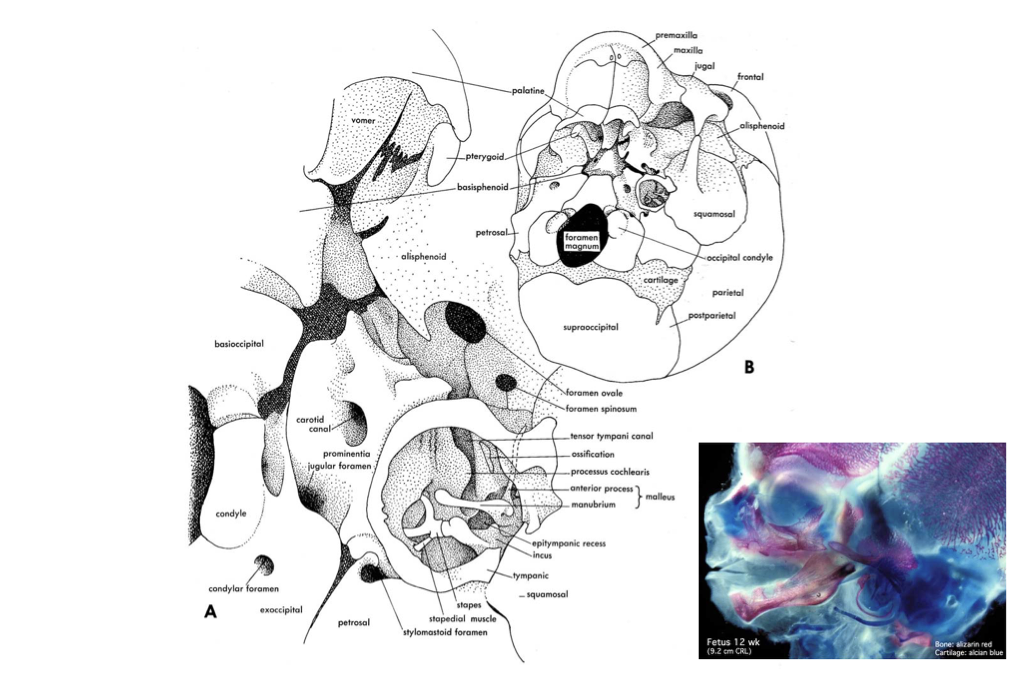
Figure 4.15. Illustration from Jollie (1962) under CC0 public domain; fetal head image acquired by Virginia Diewert and curated by Mark Hill of UNSW Embryology.


{kind=link}
{kind=link}
{kind=link}